Станислав Дробышевский Шанс для приматов Главы из книги "Достающее звено"
О книге
Главы из книги известного российского антрополога, кандидата биологических наук Станислава Владимировича Дробышевского. Полный текст можно прочитать на сайте Антропогенез. ru — http://antropogenez.ru/zveno/.
На фоне того невежества, граничащего с откровенным мракобесием, которое затопило российское телевидение и другие СМИ в последние годы, очень остро чувствуется нехватка интересных умных книг, рассказывающих об эволюции человека; раскрывающих современное состояние исследований в этой области. Кроме книг Александра Маркова "Обезьяны, кости и гены" и "Обезьяны, нейроны и душа", по этой теме, пожалуй, и почитать нечего.
Остается Интернет, но не каждый найдет достаточно времени, сил и терпения "бродить" по специализированным сайтам (даже если это такой замечательный сайт, как http://antropogenez.ru/). Не дожидаясь, когда книга Станислава Дробышевского будет напечатана (если она когда-нибудь вообще будет напечатана), предлагаем Вашему вниманию главы из нее. Думается, такая легко написанная и хорошо иллюстрированная книга будет интересна многим.
Разделы[1] книги "Достающее звено":
* Методы познания, антропогенез и палеоантропология.
* Что отличает нас от обезьян?
* Кто такие приматы. Родословная и ископаемые формы.
* Гоминиды: австралопитековые, ранние и поздние Homo.
* Исследования пропорций тела древних людей
* Археология нижнего палеолита
* Археология верхнего палеолита
* Расовая изменчивость
* Проблемы классификации гоминид
* Центр и периферия. О «протоморфности» и «недифференцированности»
* Шанс для приматов
* Палеоантропология Австралии и Океании
Введение Древнейшие млекопитающие и их классификация
Отличия человека от моржей, львов и даже кротов для большинства людей вполне очевидны. Вопрос об отличии от ежей может показаться странным, но это как поглядеть. С палеонтологической точки зрения вопрос актуален, животрепещущ и совсем не прост. Казалось бы, отличия вполне очевидны, однако не во всех аспектах.
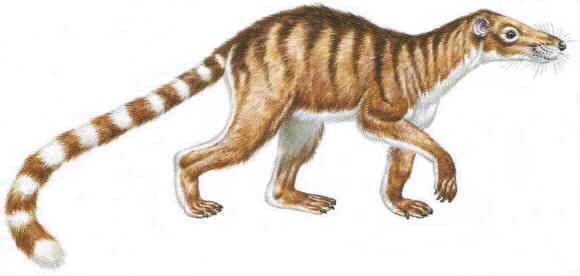
Приматы — удивительно разнообразная группа животных. У нормального человека при слове "примат" в мыслях, наверное, появляется облик макаки, шимпанзе или мартышки из мультфильма. Ассоциация с лемуром, думается, возникает лишь у статистически незначимого меньшинства. Однако, немалую часть своей истории приматы были преимущественно лемуроподобными существами, да и в настоящее время примерно треть видов относятся к ним. Впрочем, мультфильм "Мадагаскар" в последнее время мог выправить статистику в этом отношении. Но вот долгопятов, надо думать, вспоминают уж совсем редкостные чудаки. А ведь палеонтологически долгопятоподобные приматы составляли огромную и очень важную группу приматов. А от долгопятов и лемуров недалеко и до плезиадаписов. А от плезадаписов до ежей — рукой подать…

Морганукодон — одно из древнейших млекопитающих (поздний триас, более 200 млн. лет назад).
Древнейшие млекопитающие типа Morganucodonta, их позднетриасовые предки — прото-млекопитающие типа Adelobasileus cromptoni и Sinoconodon rigneyi, равно как и предки этих предков — циногнатовые Tritylodontidae — не отличались разнообразием внешности и образа жизни, которые ограничивались стилем землеройки (хотя большинство тритиллодонтов были растительноядными). С ранней юры среди Docodonta и других млекопитающих появились довольно разнообразные формы, однако подавляющее большинство их не имеют к современным зверям прямого отношения.

Останки верхнемеолового млекопитающего Cimolestes
Общими предками и насекомоядных, и приматов, и рукокрылых, и хищных, и панголинов могли быть примитивные верхнемеловые звери вроде Cimolestes.
Они имеют настолько "обобщённое" строение, что никак не помещаются в формальные классификационные схемы, зато годятся на роль всеобщих пращуров.
Предложено выделение отряда цимолестов Cimolesta, объединяющего массу верхнемеловых и раннепалеоценовых зверей, включая разномастные подотряды пантолестов Pantolesta, дидельфодонтов Didelphodonta, тениодонтов Taeniodonta, апатотериев Apatotheria, пантодонтов Pantodonta, тиллодонтов Tillodontia, палеориктид Palaeoryctidae и даже, возможно, панголинов Pholidota с заламдолестесами Zalambdalestidae, но рамки такого отряда кажутся чересчур резиновыми; сам Cimolestes иногда включается и в палеориктид Palaeoryctidae, и в дидельфодонтов Didelphodonta. Видимо, основной бум возникновения новых отрядов пришёлся на верхний мел, тем более что в начале палеоцена отряды становятся более-менее различимы. Впрочем, и тогда разница между насекомоядными, первыми копытными и хищными вовсе не всегда очевидна. Иллюстрацией могут служить дидимокониды Didymoconidae, относившиеся к креодонтам Creodonta или мезонихиям Mesonychia, а ныне выделенные в собственный отряд дидимоконид Didymoconida (Лопатин, 2001). Последовательность возникновения отрядов остаётся невыясненной по палеонтологическим остаткам. Тут могут помочь данные генетики.
Известно, что классификации, построенные по генетическим данным, резко отличаются от "морфологических". Например, по генетическим данным ежиные с землеройковыми и рукокрылые попадают в разные подразделения группы лавразиатериев Laurasiatheria, приматы с шерстокрылами и тупайи — в разные ветви эуархантоглиресов Euarchontoglires (или в одну — в зависимости от схемы), а тенреки с прыгунчиками — в афротериев Afrotheria; на высоком уровне лавразиатерии с эуархантоглиресами объединяются в бореоэвтериев Boreoeutheria и противопоставляются афротериям с неполнозубыми Xenarthra. По морфологии же все они до крайности схожи и вполне могут быть включены в единую группировку. На первый взгляд, разница капитальна, однако на второй — парадокс легко разрешим. В меловом периоде среди примитивных плацентарных дифференциация ещё не зашла слишком далеко, чтобы можно было различать их на надсемейственном уровне; однако, некий набор мутаций в разных группах различался, не влияя, впрочем, на внешний вид, принципиальные особенности морфологии и этологии; с тех пор этот набор незначимых генетических отличий ещё заметно усилился. В итоге, мы имеем несколько современных линий, примитивные представители которых сохранили морфологический план предков, но имеют генетическое расхождение, восходящее к самым основам плацентарных.
В сущности, разница "морфологических" и "генетических" схем — это разница "горизонтальной" и "вертикальной" — кладистической — таксономии. У обоих подходов есть плюсы и минусы. Морфологический — единственный применимый в палеонтологии, но резко ограничен неполнотой палеонтологической летописи и случаями конвергенции и резкой специализации. Генетический подход даёт нам представление о последовательности расхождения филогенетических линий, что лишь редко и с трудом удаётся определить по ископаемым находкам. Однако, генетический подход на современном уровне не даёт представления о сущности и масштабе находимых генетический отличий и пока мало надёжен для определения времени расхождения эволюционных линий. "Вертикальная" систематика опирается только на точки дивергенции, и, в сущности, зависима от времени: давно разошедшиеся группы считаются резко различными, даже если за миллионы лет они практически не поменялись, тогда как недавно дивергировавшие таксоны не имеют шансов получить высокий ранг, сколь бы ароморфными не были их достижения. Те незначительные мутации, которые в настоящее время различают какие-нибудь виды одного рода, в далёком будущем могут быть расценены как значимые для выделения отрядов. То есть сейчас мы считаем их видами одного рода, а систематики далёкого будущего будут вынуждены числить их в разных отрядах, даже если за это время у них не появится существенных отличий в строении и поведении. Или же, в гипотетическом пределе два древних детёныша из одного помёта, у одного из которых появилась некая мутация, должны быть с точки зрения "генетической" кладистики относены к разным отрядам, очень древних — к разным классам, а ОЧЕНЬ древних — к разным типам. У "горизонтальной" систематики свой труднопреодолимый минус — вечная проблема выбора приоритетных для систематики признаков.
Компромиссный путь пока не выработан; видимо, он должен быть "двух-" или даже "трёхмерным" — учитывать и время расхождения линий, и суть появляющихся отличий.

Слева — череп апатемиида Sinclairella; справа — кисть апатемиида Labidolemur kayi.
Позднемеловые и палеоценовые звери эволюционировали странными путями (Кэрролл, 1993а, б; Основы палеонтологии, 1962). Внешне они были большей частью весьма сходны — мелкие землеройкоподобные создания с остренькой мордочкой, пятипалыми лапками и длинным хвостом. Сугубо внешностью сходство не ограничивалось. Хотя выделяют несколько отрядов палеоценовых млекопитающих, различия их бывают слабоуловимы даже для специалистов. Дополнительной сложностью является то, что большей частью от этих зверюшек сохранились лишь зубы. А строение зубов, понятно, сильно привязано к питанию, а питание, понятно, у всех было, во-первых, схожим, то есть отсутствовали строгие различия между отрядами, а во-вторых, могло и меняться от вида к виду, то есть внутриотрядное разнообразие было большим. Лучше могли бы работать признаки основания черепа — например, набор костей в составе слуховой капсулы, — но основание черепа сохраняется плохо, у зверюшек размером с мышь и подавно, и известно для небольшого количества древних видов. Такая ситуация приводит к тому, что известен целый ряд видов, родов, семейств и даже более крупных таксонов, "зависающих" где-то между отрядами.
Например, апатемииды Apatemyidae, выделяемые обычно в собственный отряд Apatotheria, зависают между насекомоядными, плезиадаписовыми и приматами; анагалиды Anagalidae вроде бы очень похожи на тупайй (которые сами промежуточны между насекомоядными и приматами), но тоже имеют специфику, позволяющую считать их самостоятельным отрядом Anagalida; Amphilemuridae включались в приматов, но ныне зачислены в надсемейство или отряд ежиных Erinacoidea или Erinaceomorpha. Как минимум два семейства шерстокрыловых — Mixodectidae и Placentidentidae — имеют необычайное сходство с насекомоядными. Целое надсемейство микросиопоидов Microsyopoidea имеет спорный статус, его коренное семейство Microsyopidae от схемы к схеме плавает между грызунами, насекомоядными, шерстокрылами и приматами. Picrodontidae раньше считались насекомоядными, теперь — плезиадапиформами. Adapisoriculidae гуляют от сумчатых и насекомоядных до тупайй и плезиадапиформов.

Hadrocodium — череп и реконструкция
Древнейшие млекопитающие или их непосредственные предки были крошечными животными, примером чему служит Hadrocodium wui из ранней юры Китая: они достигал всего 3,2 см в длину и весил 2 грамма (Luo et al., 2001). Правда, ещё более древние позднетриасовые млекопитающие типа Morganucodon или Megazostrodon (иногда объединяемые в отряд Morganucodonta) были всё же побольше — аж целых 10 см, но тоже входили в весовую категорию землероек. Понятно, что сохранилось от них немного, кости столь малых созданий могут не очень явно отражать какие-то особенности строения мускулатуры, да и оценить таксономические различия на таком материале крайне сложно.

Скелет среднеэоценового адапида Smilodectes
Один из важнейших для систематики млекопитающих признаков — строение слуховой капсулы. Строение слуховой капсулы может варьировать в пределах одного отряда, но в общем и целом отражает родство. У тупайй слуховая капсула образована энтотимпаником — внутренней барабанной костью; такое же строение имеется у лептиктид, некоторых насекомоядных, некоторых прыгунчиковых, анагалид, рукокрылых и множества более далёких групп. У шерстокрылов слуховая капсула образована, кроме энтотимпаника, ещё и эктотимпаником — наружной барабанной костью, что встречается у грызунов, зайцеообразных и разного рода копытных. У приматов её основным элементом является каменистая кость — редкостный вариант, имеющийся только у некоторых ежиных, тенреков и прыгунчиковых (у других ежиных, тенреков и прыгунчиковых встречаются и другие варианты строения, в том числе уникальные). Таким образом, приматы отличаются от подавляющего числа других отрядов, включая тупайй и шерстокрылов. Свидетельствуют ли эти факты в пользу особо близкого родства приматов и насекомоядных и отдаления их от тупайй и шерстокрылов? Скорее всего — нет. У некоторых ранних групп приматообразных животных окостеневшая слуховая капсула могла отсутствовать или не прирастать к черепу, в этом уличены Ignacius, Phenacolemur и Microsyops из плезиадапиформов, а вот адапиды — например, Pronycticebus — уже имели каменистую слуховую капсулу, сросшуюся с основанием черепа. То же прослеживается в ранней эволюции ежиных — у древнейших форм капсула не окостеневала. Таким образом, окостенение капсулы появилось позже сложения таксономических групп и не может явно говорить о их родстве.

Скелет плезиадаписа (поздний палеоцен)
У многих групп палеоценовых и эоценовых млекопитающих проявилась тенденция к развитию длинных долотоподобных передних зубов, далеко выступающих вперёд, с постоянным ростом и редуцированной эмалью. Этим отличались многобугорчатые, некоторые сумчатые, залямбдалестиды, тиллодонты, поздние тениодонты, апатемииды, некоторые ископаемые землеройки и ежи, грызуны, зайцеообразные, даманы, паромомиоиды, микросиопоиды, плезиадапиформы и плезиопитециды, не говоря о совсем уж экзотичных хоботных, пиротериях, десмостилиях и нотоунгулятах. С одной стороны, эта особенность сама по себе мало о чём говорит, так как сопровождается самыми разными вариациями строения прочих зубов и возникала по разным поводам. Склонность резцов к увеличению обеспечивалась за счёт разных мутаций, и в разных группах увеличивались разные резцы (у тениодонтов вместо резцов ту же форму приняли вообще клыки). Показательно, что, несмотря на неоднократное возникновение такой зубной системы среди разных линий приматов, в наши дни её имеет лишь один вид — мадагаскарская руконожка. Вероятно, остальные подобные линии не выдержали конкуренции с грызунами, а руконожку спасла её крайняя специфика и изоляция (впрочем, гигантской руконожке, вымершей около тысячи лет назад, и это не помогло). Несмотря на то, что относительный размер мозга у руконожки — один из самых больших среди полуобезьян, особой гениальности за ней пока не замечено.
Кроме строения зубов и слуховой капсулы, приматы отличаются от насекомоядных крупными размерами глаз. Насекомоядные вообще видят плохо, глаза у них крошечные. Приматы видят хорошо и имеют большие глаза. Эти различия возникли, вероятно, очень рано, ещё на стадии плезиадапиформов и были, вероятно, связаны с образом жизни — в лесной подстилке для насекомоядных и на деревьях и кустарниках — для приматов. Соответственно, обонятельный орган у насекомоядных велик, а у приматов — мал. Впрочем, прыгунчики имеют большие глаза, а Palaechton из плезиадапиформов имел маленькие, так что и этот признак не идеален.
Таким образом, практически нет каких-то универсальных и к тому же легко проверяемых признаков, которые бы помогли оценить эволюционную близость групп млекопитающих.
Поэтому палеонтологам приходится хитрить и мудрствовать, чтобы выявить преемственность или, напротив, конвергенцию сходных форм.
Древнейшие насекомоядные, землеройки и ежи
Древнейшие насекомоядные — Paranyctoides — известны из середины верхнего мела — чуть более раннего времени, чем древнейшие приматоподобные существа. Впрочем, как обычно бывает, таксономическая принадлежность этих ранних форм спорна. Они равно могут быть сближены с ежеподобными и землеройкоподобными. Достоверные насекомоядные в узком смысле Eulipotyphla появляются в палеоцене.

Миоценовый ёж Brachyerix
Тогда в Америке, Европе и Северной Африке широко были распространены палеориктиды Palaeoryctidae — землеройкоподобные животные, но они, как и землеройки Soricidae, с самого начала были привязаны к земле и оказались достаточно специализированными роющими животными, к тому же особо-насекомоядными, судя по очень высоким и острым бугоркам зубов (что в будущем сделало из них хищников — Creodonta и Carnivora). В среднем и позднем палеоцене уже имеются представители семейства ежовых Erinaceidae, например, североамериканский Mackennatherium и германский Adunator.
Ежовые, таким образом, — чуть ли не самое древнее из современных семейств млекопитающих, исключая разве что броненосцев Dasypodidae.
Неудивительно, что они сохранили массу примитивных черт, а потому плохо отличимы от прочих древних зверей и имеют множество сходств с совершенно разными современными млекопитающими.

Челюсть ископаемой землеройки Limnoecus (миоцен)
Никтитериды Nyctitheriidae — существа, возможно, не только землеройкоподобные, но и близкородственные собственно землеройкам Soricidae, известны из палеогена. Любопытно, что первоначально они были древесными, в противоположность современным родичам и, возможно, потомкам — землеройкам Soricidae и кротам Talpidae. К среднему эоцену насекомоядные имели уже почти современный облик и вели нынешний образ жизни. С деревьев они были почти полностью вытеснены более развитыми приматами.
Впрочем, вытеснялись они, возможно, не только вниз — в почву, но и вверх — в воздух, ведь часть никтитерид, вероятно, дала летучих мышей.
Зубы никтитерид и рукокрылых почти неотличимы и не исключено, что часть палеоценовых родов, известных лишь по зубам, относятся к древнейшим летучим мышам.

Череп лептиктида Palaeictops bicuspis (ранний эоцен, Северная Америка)
Между насекомоядными и шерстокрыловыми промежуточны палеоценовые Mixodectidae, а Г. Осборн считал их грызунами. Предками миксодектид могли бы быть палеориктиды, ежиные или их близкие родичи (Szalay, 1969). Близки к предкам приматов лептиктиды Leptictida — особый отряд палеогеновых зверей, — хотя даже самые ранние их представители — Gypsonictops из верхнего мела — оказываются более специализированными, чем миксодектиды. Лептиктиды были хищными и явно специализировались к передвижению на задних ногах, причём не только пряжками, но и бегом (Leptictidium), некоторые стали внешне несколько похожи на современных прыгунчиков, а размеры их колебались от мышиных до длины в полметра и даже больше. Строение лептиктид, кстати, во многом похожих на ежиных, с одной стороны примитивно, с другой — у них имеются явные черты специализации; это позволяет говорить о близости лептиктид ко всем примитивным отрядам млекопитающих, включая приматов, но не позволяет утверждать, что лептиктиды были их прямыми предками. Имеются и другие верхнемеловые млекопитающие, имеющие в целом примитивное строение, но с элементами специализации, например, Procerberus. Как итог, их систематика остаётся крайне спорной.

Скелет апатемиида Heterohyus nanus с сохранившимися мягкими тканями
Апатемииды Apatemyidae — одна из групп неопределённого положения; их относили и к насекомоядным, и к плезиадапиформам, и выделяли в собственный отряд апатотериев Apatotheria, говорилось о близости апатемиид к грызунам, копытным, хищным, тениодонтам, тиллодонтам, кондиляртрам и, конечно, приматам. Впрочем, их зубы оказались слишком специализированы уже к началу палеоцена, чтобы можно было выводить из апатемиид кого-либо. Сильно увеличенные передние резцы — особенно нижние — делали их похожими на плезиадапиформов, грызунов и кускусов, близки они были и по стилю жизни, но сильно уступали большинству конкурентов в размерах, так как были величиной с мышь. Апатемиид отличала очень массивная голова, вытянутые тонкие пальцы и крайне длинный хвост. Жили они на деревьях.

Полосатый поссум Dactylopsila trivirgata (Новая Гвинея)
По всей видимости, апатемииды питались преимущественно насекомыми, которых добывали из-под коры деревьев подобно руконожкам и новогвинейским полосатым поссумам Dactylopsila — простукивая стволы, прогрызая в них дырочку мощными зубами и доставая добычу удлинёнными вторым и третьим пальцами. Показательно, что ни на Мадагаскаре, ни на Новой Гвинее нет дятлов. Апатемииды известны со среднего палеоцена Европы и Северной Америки;
поздние виды дотянули до среднего олигоцена, но, по всей видимости, проиграли эволюционную гонку приматам, грызунам и — в особенности — дятлам (Koenigswald et Schierning, 1987).
Древнейшие дятлы, хотя ещё и без своей знаменитой долбёжной адаптации, известны из того же среднеэоценового местонахождения Мессель в Германии, что и апатемиид Heterohyus; отверстия, похожие на следы работы дятлов, найдены в стволах эоценового леса в Аризоне. Поздние апатемииды увеличились в размерах и, вероятно, пытались перейти на новые источники пищи, но дятлы победили. Ставка на специализацию, как обычно, оказалась роковой. Аналогичная адаптация, возможно, возникла у крупного плезиадапида Chiromyoides в позднем палеоцене Северной Америки и Европы. Любопытно, что предок руконожки Plesiopithecus teras с характерной специализацией зубов известен из верхнего эоцена Египта, но и сюда добрались коварные дятлы, так что руконожка нашла убежище лишь на далёком лесистом острове.
Все насекомоядные в широком смысле имеют очень слабое развитие мозга. Интеллект у ежа понятно какой.
Да и каким ему быть, коли масса мозга у него — чуть больше трёх грамм, извилин на нём нет, а большая часть отвечает за обоняние?
Ежу понятно, что извилинами с таким мозгом не пошевелишь — извилин-то нет! Малые размеры тела, правда, обеспечивают землеройкам рекордные показатели относительной массы мозга, но они же приводят к необходимости высочайшего метаболизма — опять же рекордного для млекопитающих. Необходимость согревать махонькое тельце не оставляет энергии и возможностей для сколь-либо существенного интеллекта и, кроме прочего, сокращает продолжительность жизни. Ежи, правда, зимой превращаются фактически в холоднокровных животных, но спячка ещё в меньшей степени способствует разумности.
Древнейшие грызуны
Грызуны возникли, вероятно, как группа роющих существ, питающихся твёрдыми растительными кормами, по крайней мере многие древнейшие представители вели именно такой образ жизни. Часть даже самых ранних верхнепалеоценовых и нижнеэоценовых родов — Paramys и Ischyromys — имели белкоподобный облик и, возможно, лазали по деревьям, но конкуренция со стороны плезиадаписовых — тоже древолазящих и грызуноподобных в строении черепа, но более интеллектуальных — очевидно, была слишком велика.

Древнейший грызун Ischyromys oweni (ранний эоцен)
Поэтому, несмотря на то, что в последующем — где-то с верхнего эоцена и после — многие грызуны перешли к древолазанию и питанию фруктами, а многие стали не прочь съесть и чего животного (летяги — страшные враги всех певчих птиц), грызуны не смогли выйти на новый эволюционный уровень. К этому времени в тропиках приматы уже прочно заняли соответствующую экологическую нишу, а вне тропиков, видимо, не хватало фруктов и развесистых деревьев, так что у белок, сонь и древесных дикобразов не осталось шансов стать разумными. Впрочем, не исключено, что спустя миллионы лет грызуны отыгрались и "сказали таки своё веское слово" в вымирании парантропов, но это можно считать лишь пакостной местью неразумных созданий, но никак не заявкой на эволюционно-интеллектуальное превосходство.
Однако, примитивность в сочетании с неимоверными темпами видообразования — максимальными среди млекопитающих — оставляют грызунам надежду на разумное будущее.

Череп Spalax leucodon (малый слепыш)
Вероятно, грызуны начали эволюционную гонку несколько позже приматоморфов. Мелкие размеры (отдельным видам — Phoberomys pattersoni из верхнего миоцена Венесуэлы и Josephoartigasia monesi из плиоцена Уругвая — удалось достичь размеров очень крупной коровы или маленького носорога, но исключения — одно из основных свойств живой природы, да и большими мозгами такие мегакрысы похвастаться не могли (Sánchez-Villagra et al., 2003; Rinderknecht et Blanco, 2008)), преимущественная растительноядность, значительная морфологическая специализация, малая продолжительность жизни, большое количество детёнышей — достаточные причины для сохранения простого строения мозга и слабой социальности. Даже самые социальные суслики и голые землекопы особым интеллектом не блещут, простота добывания корма и мощный пресс хищников способны задавить всякие проблески разума. Грызуны смогли вытеснить многобугорчатых Multituberculata (которые вымерли как раз в конце эоцена — одновременно с расцветом грызунов), но "приматный" путь развития им был уже заказан.
И это при том, что эволюционный корень приматов и грызунов был един!
Древнейшие грызуны Rodentia известны из позднего палеоцена Северной Америки, а примитивнейшие — из нижнего эоцена Азии; в Азии в верхнем палеоцене тоже имеется несколько "грызуноподобных" видов, так что возникнуть отряд мог и там, и там. Эти архаичные формы очень схожи с азиатскими анагалидами Anagalida. Получается интересная ситуация: анагалиды признаны неродственными приматам, приматы — близки к грызунам, а грызуны, тем не менее — потомки анагалид. Путаница была распутана новейшими исследованиями, в которых было показано, что сходство грызунов с анагалидами конвергентно. В настоящее время в качестве предкового для грызунов и зайцеобразных (или же только зайцеобразных) называется эндемичный азиатский отряд миксодонтов Mixodontia и даже конкретнее — мимотониды Mimotonida и эвримилоиды Eurymyloidea (Лопатин, 2004), известные с раннего палеоцена. Любопытно, что древнейшие приматоморфы связывают Северную Америку с Европой, а грызуны — Северную Америку с Азией; вероятно, это связано с распространением в области с наименьшим сопротивлением со стороны других отрядов. Не значит ли это, что в палеоцене и эоцене в Азии кто-то составлял приматам серьёзную конкуренцию? Впрочем, дело может быть просто в благоприятности экологических условий. Может, в Азии в палеоцене было меньше густых лесов, где могли бы жить приматы?
Как итог, грызуны имели с приматами одних предков в верхнем мелу, но в палеоцене уже заметно разошлись — как экологически и морфологически, так и по эволюционной судьбе.
Почему зайцы не стали приматами?
Близки к грызунам зайцеобразные Lagomorpha. Препятствия на пути "приматизации" зайцеобразных были теми же, что и для грызунов, хотя и с любопытной поправкой. Строение челюстей, тип прикуса и механика пережёвывания пищи зайцеобразных — с боковыми движениями — позволяют каждому виду осваивать широкий спектр экологических условий и огромные ареалы без особой морфологической перестройки.

Череп ископаемого зайца Palaeolagus
Грызуны же специализируются именно на морфологическом уровне. Поэтому современные зайцеобразные насчитывают от силы полсотни видов, тогда как грызуны — несколько сотен родов и около 1700 видов. Таким образом, зайцеобразные с самого начала своей эволюции ступили на путь "специализации к неспециализации", столь часто постулируемый как особо прогрессивный и якобы давший приматам невероятные эволюционные преимущества. Однако, зайцеобразные "слишком рано" "вышли в саванну" — специализировались к наземному бегу и, хотя древесные зайцы тоже в природе существуют, так и не развили сколь-либо выраженной подвижности и цепкости "рук". Зацикленность на растительной пище у зайцев ещё бóльшая, чем у грызунов, так что особых поводов для увеличения мозга и усложнения поведения в их истории не возникало. Пищухи Ochotonidae достаточно социальны, но не настолько, чтобы их можно было сравнивать даже с лемурами.
Да и возникли зайцеообразные слишком поздно — лишь в среднем эоцене, так что шансов составить конкуренцию приматам у них уже не оставалось.
А копытные?
Ещё одна крайность — копытные травоядные Ungulata. Среди палеогеновых млекопитающих особым разнообразием отличалась группа кондиляртр — Condylarthra, возникшая в самом начале палеоцена. Древнейший — Kharmerungulatum vanvaleni — найден в позднем мелу Индии (Prasad etal., 2007). Несколько лучше представлены северноамериканские роды кондиляртр Protungulatum, Oxyprimus, Baioconodon и Mimatuta. Protungulatum gorgun был найден в слоях, содержащих зубы динозавров, а потому обычно упоминается как позднемеловой.

Кондиляртр Hyopsodus (эоцен)
Выяснилось, однако, что течением реки слои позднего мела и раннего палеоцена были перемешаны; Protungulatum с наибольшей вероятностью является раннепалеоценовым. Как и для всех прочих плацентарных, корни кондиляртр теряются в густом тумане границы мела и палеоцена; в качестве предков назывались желестиды Zhelestidae — мелкие зверьки позднего мела Средней Азии, Европы и Северной Америки, — но родство этих групп поставлено под основательное сомнение (Archibald et Averianov, 2012; Wible et al., 2007). Сами кондиляртры с наибольшей вероятностью являются сборной группой, но известные черты её представителей столь плохо различимы, что практически невозможно адекватно разделить её на части (Wible et al., 2007).

Hyopsodus (ранний — средний эоцен, возможно поздний палеоцен)
Среди кондяляртр можно найти формы, весьма напоминающие приматов. Их зубы тоже имеют притуплённые бугорки, более-менее прямоугольную форму коронок и прочие тонкости, схожие с признаками приматов. Неспроста статус некоторых животных долгое время оставался или до сих пор остаётся под вопросом: кондиляртры это или приматы? Примером могут служить Decoredon anhuiensis из палеоцена Китая, Hyopsodus и Promioclaenus из палеоцена и эоцена Северной Америки. Первоначально кондиляртры были стопоходящими всеядными животными, причём в диете немалую роль играли плоды и листья — как и у приматов, а сами они могли лазать по деревьям. Неудивительно, что некоторые тенденции эволюции самых ранних — нижне- и среднепалеоценовых — представителей группы весьма напоминали то же у приматов. Например, бугорки на молярах тоже стали понижаться, зубы стали закругляться и вытягиваться. Однако, в дальнейшем кондиляртры пошли большей частью по пути приспособления к наземному бегу и питанию жёсткой растительной пищей, дав, в числе прочих, парно- и непарнокопытных Artiodactyla и Perissodactyla. Посему морда их стала удлиняться, а не укорачиваться, как это было у приматов, жевательные зубы резко увеличились, выросло и число бугорков на них, конечности стали менее гибкими и полупальцеходящими, а первый палец не только не получил тенденции к противопоставлению, но вообще стал редуцироваться, вместе с пятым. Когти преобразовались не в ногти, а в копыта.

Примитивное копытное Agriochoerus antiquus
Предполагалось, что некоторые примитивнейшие копытные с когтями на пальцах — эоценовые североамериканские агриохериды Agriochoeridae — могли даже лазать по деревьям, хотя вряд ли очень ловко, так что конкуренции на этом поприще с уже существовавшими тогда приматами они, конечно, выдержать не могли. Показательно, что древнейшие копытные отличались примитивным строением черепа и зубов, но имели довольно специализированные конечности.
Учитывая, что бóльшую часть сведений о древних млекопитающих мы извлекаем из зубов, этот факт заставляет задуматься.
Кондиляртр Chriacus arctocyonid (ранний палеоцен — ранний эоцен, Северная Америка)
Одним из "звоночков", которые могли определить будущую судьбу кондиляртр, является приспособленность самых ранних представителей не просто к жизни на земле, но к рытью. Хорошим примером может служить род Chriacus, разные виды которого жили в Северной Америке на протяжении всего палеоцена и в начале эоцена. Chriacus имел гибкие конечности с пятью пальцами, увенчанными когтями, он мог бегать, лазать по деревьям, ел всё подряд — фрукты, насекомых, мелких животных, его нижние резцы даже приобрели вид "зубной щётки" для чистки шерсти — как у лемуров. Но его передние конечности явно предназначены иногда копать, — и этим кондиляртры отличаются от приматов. Кондиляртры были универсальнее, приматы — специализированнее, а потому первые не смогли вытеснить последних на деревьях, а последние не могли спуститься на землю. Некоторые кондиляртры довольно долго вели соревнование с приматоморфами за жизнь на деревьях: белкоподобные гиопсодонтиды Hyopsodontidae появились на границе палеоцена и эоцена — тогда же, когда настоящие приматы и грызуны. Те, другие и третьи были очень схожи экологически и, как итог, имели массу параллелей в морфологии. Однако приматы и грызуны имели более длинную историю становления древесности, а землю уже захватили потомки кондиляртр, так что гиопсодонтидам не оставалось ничего, кроме вымирания (хотя поначалу они, как всякие малоспециализированные формы, были крайне многочисленны и часто представляют большинство млекопитающих в местонахождениях).

Останки пантодонта Archaeolambda planicanina
Большинство потомков кондиляртр стали специализированными растительноядными копытными животными, однако некоторые и после "имели шанс" вступить на "приматный путь". К примеру, среди пантодонтов Pantodonta монгольская нижнеэоценовая Archaeolambda planicanina, насколько это видно из её зубов, была всеядно-насекомоядной, приматы тогда в Монголии почти отсутствовали (достоверно известен лишь один вид Altanius orlovi), так что некоторые пантодонты могли занимать их экологическую нишу; впрочем, вряд ли археолямбда была очень похожа на приматов, учитывая внешний вид других пантодонтов. Впрочем, пантодонты не смогли превзойти даже представителей южноамериканской фауны, известных своей низкой конкурентноспособностью — в раннем палеоцене Южной Америки жил пантодонт Alcidedorbignya inopinata, а в последующем эта группа там исчезла (одновременно сей факт говорит о наличии хотя бы ограниченной связи между материками в раннем палеоцене, так что теоретически приматоморфы могли заселиться в Южную Америку уже тогда; а может — тоже заселились, но после исчезли? Вдруг новейшие изыскания откроют нам южноамериканских палеоценовых плезиадапиформов?).
Как выглядели и жили некоторые ранние растительноядные, можно представить, наблюдая современных даманов Hyracoidea, хотя современные даманы, без сомнения, достаточно специализированы и не могут считаться "живыми ископаемыми". Часть их признаков отдалённо напоминает вариант приматов, например, стопо- или полустопохождение, плоские ногти на пальцах (коготь сохраняется на первом пальце задней ноги — точь-в-точь как у лемуров), способность лазить по деревьям (впрочем, за счёт влажности подушечек пальцев, а не их хватательной способности). Однако, всё же подавляющая часть их черт явно сближается с более крупными растительноядными; явного родства с приматами у даманов, конечно, нет.

Диноцерас (Dinoceras).
Переход к растительноядности независимо повторили несколько групп млекопитающих. Например, ранние среднепалеоценовые диноцераты Dinocerata были похожи на ранних кондиляртр и креодонтов, но скоро превратились в огромных носорогоподобных зверей с рогами и копытами. Такая же судьба ожидала и многих других зверей. Эволюция их совершалась, как правило, очень быстро, даже самые ранние представители уже имеют все главные признаки своей группы. Впрочем, некоторые смогли сохранить некоторые первоначальные черты вплоть до современности. Хорошим примером могут служить свиньи Suina. Свинообразными были самые первые парнокопытные и их примитивные зубы — низкокоронковые бугорчатые — во многом напоминают, с одной стороны, зубы приматов, с другой — сохраняют общий план у современных свиней. В немалой степени этому способствует, конечно, всеядный характер питания. Масса примитивных признаков, кстати, сохраняется у них и в строении конечностей. Фактически, свиней можно рассматривать как эволюционно очень продвинутый экологический аналог наземных приматов (наше далёкое будущее?).
Все наземные растительноядные невыгодно отличаются от приматов низким интеллектом и упрощённо-механизированными конечностями, движущимися лишь в одной продольной плоскости и потерявшими всякие шансы стать хватательными.
Низкокалорийность растительной пищи приводит к усложнению и специализации пищеварительной системы, оттягивающей на себя энергетические и эволюционные акценты. С другой стороны, корневища, трава и листья не прячутся, не убегают и активно не сопротивляются. Добыть их не сложно, а цвет почти всегда зелёный, что ведёт к редукции цветного зрения. Всё это в совокупности приводит к малой интеллектуальности копытных, вошедшей в массу поговорок и ругательств.
Хищники — тоже не приматы?
Обратная крайность — хищники. Хищный образ жизни вели предки млекопитающих ещё до появления этой группы. Древнейшие млекопитающие все были хищниками в широком смысле слова. Впрочем, поеданием червячков и букашек дело не ограничивалось, о чём недвусмысленно свидетельствуют кости пситтаккозавра в желудке триконодонта Repenomamus robustus из нижнего мела Китая (Hu et al., 2005). А ведь родственный ему Repenomamus giganticus был вдвое больше — около метра в длину.

Repenomamus robustus поедает пситтаккозавра (нижний мел, Китай)
Хищники уже с самых ранних форм — нижнепалеоценовых кондиляртр арктоционид Arctocyonidae и верхнепалеоценовых креодонтов Creodonta — достаточно специализированы, но в их строении проглядывает более древнее состояние: когти не очень острые, зубы не имеют хищнического лезвия и в целом похожи на зубы насекомоядных и приматов, в скелете имеется хорошо развитая ключица, конечности стопоходящие, многие хорошо лазали по деревьям. Вероятно, они были ещё не абсолютно хищными, а всеядными. Из родственников современных хищных уже в нижнем палеоцене виверравиды Viverravidae были заметно специализированными к питанию насекомыми. Вместе с тем, у среднепалеоценовых миацид Miacidae на фоне усиления хищнического комплекса имелся такой почти "приматный" признак, как умеренное противопоставление большого пальца. Виверравиды и миациды в настоящее время часто определяются как Miacoidea и исключаются из хищных в узком смысле слова; в середине эоцена появляются настоящие хищные современного отряда Carnivora, но преемственность всех этих групп достаточно очевидна, а экологически они не особо отличались. Таким образом, хищники в широком смысле Carnivoramorpha успели занять свою экологическую нишу, по сути дела, раньше приматов, так что можно рассуждать о том, почему приматы не стали хищнее, чем могли бы.
Конечно, гоминиды в итоге отыгрались по полной программе (все крупные и многие мелкие хищники сейчас в Красной Книге), но до этого было ещё далеко.
Впрочем, многие хищные вторично вернулись ко всеядности и даже специализированной растительноядности. Показательно, что, например, у енотовых Procyonidae в связи с этим моляры приобрели квадратную форму и притуплённые бугры, а лапки развили необычайные манипулятивные способности. Ещё больше похожи на зубы приматов моляры большой панды Ailuropoda melanoleuca.

Арктоционид Hyracolestes ermineus (нижний палеоцен, Монголия)
Причина "неприматизации" хищников видится в несоциальности всеядных форм и хищнической специализации социальных; про специализированных несоциальных можно вообще не вспоминать. Медведи Ursidae могут собираться группами при изобилии корма — например, нерестовом ходе лосося или созревании ягод, — но при этом никак не общаются между собой, а, напротив, стремятся держаться подальше друг от друга. Вероятно, так же вели себя медведеподобные по стилю жизни мезонихии Mesonychia — странные звери с огромными челюстями, маленькими мозгами и копытами на ногах, появившиеся в самом начале палеоцена в Азии и дожившие до начала олигоцена. Социальные львы — специализированные хищники на крупных копытных; впрочем, все прочие кошачьи Felidae несоциальны. Наверное, максимально приближены к "приматному идеалу" шакалы Canis и носухи Nasua — они и социальны, и всеядны. Но хищнические корни завели морфологию шакалов далеко по пути специализации, так что трудно представить, как они могут перейти к орудийной деятельности.
А вот у древесных носух с их подвижными пальцами и общительностью, наверное, неплохие шансы — не зря они так напоминают лемуров.
Настораживает лишь одно — долгие миллионы лет носухи остаются носухами и всё никак не станут чем-то бóльшим. Вероятно, препятствием на пути разумности становятся слишком длинные когти и носы? Енотовые Procyonidae появились в начале олигоцена — намного позже приматов, — может, у них всё ещё впереди?
Древнейшие приматы и многобугорчатые
Почему же именно приматы вылавировали между крайностями насекомоядных, грызунов, травоядных и хищников и пошли по пути развития интеллекта? Ведь задел у всех был один?

Зубы Purgatorius (нижний палеоцен)
Первые приматы имели менее высокие и не такие острые бугорки моляров, более мощные и широкие скуловые дуги и не столь вытянутую мордочку, нежели ежиные. Верхние зубы ранних приматов поперечно более узкие, чем у насекомоядных, есть и более специфические отличия в строении зубов, например, пониженная разница между высотой тригонида — "трёхбугорчатой" части — и талонида — "пятки" нижних моляров (Szalay, 1969). Например, древнейший приматоморф Purgatorius из верхнего мела США — вернее, единственный известный нижний моляр — определён как приматоморф именно по сочетанию квадратных очертаний с притуплённостью бугорков тригонида, точно такие же зубы известны из нижнего палеоцена (хотя у Purgatorius тригонид высокий, а талонид узкий, и имеется стилярная полка, что не позволяет однозначно определить его как примата). Все эти особенности иногда связывают с переходом с сугубо насекомоядной диеты на смешанную, включающую фрукты и листья (Szalay, 1969). Правда, судить о питании по зубам получается не всегда.
Например, у современных шерстокрылов зубы остробугорчатые, "насекомоядные", тогда как питаются эти звери листьями и плодами.
Может, меловые предки приматов имели подобное же сочетание зубов и диеты, а потому мы не узнаём их среди прочих насекомоядноподобных зверюшек?
Похожий на раннеприматный комплекс развился у многобугорчатых Multituberculata, представляющих самостоятельный подкласс Allotheria, особенно у птилодонтид Ptilodontidae, чья зубная система удивительным образом сочетала специализированность и универсальность: как ёмко охарактеризовала их В.И. Громова, выступающие вперёд "длинные резцы служили для прокалывания и прогрызания твёрдых плодов, большой задний нижний переднекоренной — для разрезания оболочек более мягких и размельчения крупных плодов, а многобугорчатые заднекоренные — для их раздавливания" (Основы палеонтологии, 1962). Такая зубная система функционально близка к совершенству и может использоваться как для питания растительными кормами, так и животными.

Череп [и реконструкция] Ptilodus (палеоцен, Северная Америка)
Некоторые многобугорчатые вели древесный образ жизни, а Ptilodus даже имел хватательный хвост и мог спускаться по деревьям вниз головой, как белки. Однако, многобугорчатые уступали приматам заметно меньшим развитием мозга. То же можно сказать о тиллодонтах Tillodontia: они были всеядны, стопоходящи, сохраняли ключицу, могли лазать по деревьям и имели потенциально хорошие эволюционные шансы. Однако, достаточно одного взгляда на их мозговую коробку — узкую, низкую, со всех сторон стиснутую жевательными мышцами, — чтобы понять причины их вымирания. Та же причина вымирания называется для целого ряда отрядов млекопитающих. Гладкий маленький мозг свёл в могилу половину ранних зверей, при том, что их образ жизни и размеры колебались от землеройкоподобных до слонопотамовидных.

Древнейшее плацентарное млекопитающее Eomaia (мел, Китай)
Другой причиной вытеснения многобугорчатых приматами и грызунами было несовершенство их онтогенеза. Судя по малому отверстию между двумя половинками таза, многобугорчатые рожали очень мелких недоразвитых детёнышей, подобно современным сумчатым (имеются и сумчатые кости, но они прямо не связаны с сумкой и имеются также у одного из древнейших плацентарных — Eomaia scansoria (Ji etal., 2002)). Сочетание с малыми размерами тела и короткой жизнью оказалось буквально убийственным: такие животные не могли быстро нарастить мозги и накопить жизненный опыт, так что их интеллектуальный уровень оставался крайне низким. К тому же челюсти многобугорчатых могли двигаться только по вертикали и вперёд-назад, а жевательные движения им были недоступны.

Череп тиллодонта Trogosus huracoides
Пока достойных конкурентов не было, всё было хорошо — так продолжалось, кстати, примерно 100 миллионов лет подряд,
— но когда приматы и грызуны достаточно развились, они задавили многобугорчатых интеллектом и обогнали их в скорости пережёвывания пищи ("хорошо пережёвывая пищу, ты помогаешь обществу!"). А ведь ещё были тениодонты Taeniodonta и тиллодонты Tillodontia— роющие животные, питавшиеся корневищами, крупные и сильные, так что конкуренция в "грызунячьей" экологической нише в палеоцене была основательной. Правда, к чести многобугорчатых, стоит отметить, что для окончательной победы приматам и грызунам понадобилось не менее 25 миллионов лет.
Почему сумчатые не преодолели "мозговой барьер"
Почти те же самые причины привели к почти повсеместному исчезновению сумчатых Metatheria. Показательно, что среди сумчатых крысовидных опоссумов-ценолестид Caenolestidae и палеогеновых полидолопид Polydolopidae развилась зубная система, очень похожая на систему многобугорчатых; видимо, они были аналогом последних в Южной Америке. В Австралии такой же вариант известен у крысиных кенгуру Aepyprymnus и карликовых поссумов Burramys.

Древнейший опоссум Alphadon (мел)
Самые примитивные сумчатые — опоссумы-дидельфиды Didelphidae и ранние дазиуроиды Dasyuroidea — экологически весьма похожи на предков приматов и древнейших приматов — едят всё подряд, хорошо лазают по деревьям, многие имеют хватательный хвост и противопоставляющийся большой палец на задней ноге (на нём даже нет когтя). Однако, приматы эволюционировали в крупных и умных животных, а опоссумы остались такими же, какими были ещё в меловом и чуть ли не юрском периоде. Конечно, сумчатые тоже не стояли на месте, из опоссумоподобных предков развились аналоги неполнозубых (сумчатый муравьед Myrmecobius fasciatus), насекомоядных (включая сумчатого крота Notoryctes), грызунов (включая сумчатых летяг Petauridae и Acrobatidae) и даже крупных хищников — размером до большого медведя или леопарда (вроде южноамериканских боргиенид Borhyaenoidea и их потомков тилакосмилид Thylacosmilidae, а также австралийских тилацинид Thylacinidae и тилаколеонид Thylacoleonidae).
Но показательно, что среди всего изобилия форм сумчатых нормальных аналогов приматов так и не возникло.
Сумчатые не дали ни бегающих хищников, ни быстрых древолазов. В Южной Америке опоссумы Didelphidae и Microbiotheriidae, а в Австралии и Меланезии коала Phascolarctos cinereus, кускусы Phalangeridae и Tarsipedidae, а равно древесные кенгуру Dendrolagus проигрывают по всем статьям даже белкам, не говоря уж о мартышках. Среди полуобезьян тоже есть медленные лори, но даже они способны двигаться быстрее древесных сумчатых. Кускусы имеют противопоставляющиеся большие пальцы, всеядны, имеют квадратные моляры — всё знакомые черты, — но на этом сходство с приматами заканчивается.

Кускус хоботноголовый Tarsipes rostratus
Хоботноголовые кускусы Tarsipes rostratus, питающиеся мёдом и цветочной пыльцой, экологически весьма напоминают некоторых мышиных лемуров, но и те, и другие являются примерами крайней специализации. Любопытно, что даже древесные кенгуру, имевшие прыгающих наземных предков, по деревьям предпочитают двигаться фактически ползком; этим сумчатые принципиально отличны от приматов, один из важнейших признаков которых — адаптация к прыганию.
Очевидно, ключевая разница сумчатых и приматов — в степени развития мозга. Что сдерживало развитие мозга аллотериев-многобугорчатых и сумчатых — вопрос. Уже упомянутое несовершенство системы вынашивания детёнышей не позволяло рожать их достаточно развитыми, чтобы уже после рождения они успели набраться ума-разума. У самых продвинутых в этом отношении сумчатых — бандикутов Peramelemorphia — имеется аллонтоисная плацента, но безворсинчатая, а трофобласта, препятствующего отторжению плода, у них нет. Посему, как и все прочие сумчатые, бандикуты рожают детёнышей фактически на стадии эмбриона.
Возможно, ещё сотня миллионов лет — и потомки бандикутов покорили бы Землю, но кто ж им даст эту сотню?
Видимо, в большом интеллекте до поры до времени не было надобности, поскольку окружение в виде ящериц и динозавров само было не чересчур интеллектуальным, конкурировать было не с кем. А после появления плацентарных было уже поздно, чему свидетельством печальная история всех сумчатых фаун на всех континентах. К примеру, конкуренция с грызунами и приматами называется в качестве основной причины вымирания сумчатых полидолопид в олигоцене. Показательно, что конкуренцию с ранними насекомоядными сумчатые вполне выдержали: ещё в нижнем палеоцене палеориктиды проникли-таки в Южную Америку (Marshall et Muizon, 1988), но потом исчезли на долгие миллионы лет (опять же — раз прошли палеориктиды, стало быть, могли пройти и плезиадапиформы, но их мы в Южной Америке не находим). Да и современные землеройки смогли заселить лишь самый север Южной Америки, южнее им успешно противостоят опоссумы, которые даже устроили "контрнаступление" на североамериканский континент.
Некоторые сумчатые, видимо, пытались преодолеть "мозговой барьер". Так, нижнепалеоценовый опоссум Pucadelphys andinus из Боливии был довольно социальным животным, о чём свидетельствует выраженный половой диморфизм и обнаружение десятков особей разного пола и возраста в одном месте — на площади одного "гнезда" (Ladevèze et al., 2011). Однако, такие продвинутые сумчатые "опередили своё время" и не получили развития, последующие виды вели одиночный образ жизни.
Скоростная эволюция: тениодонты и рукокрылые
В качестве прогрессивного признака какой-либо группы — часто приматов — иногда называется большая скорость эволюции, но это положение нуждается в категоричном уточнении.
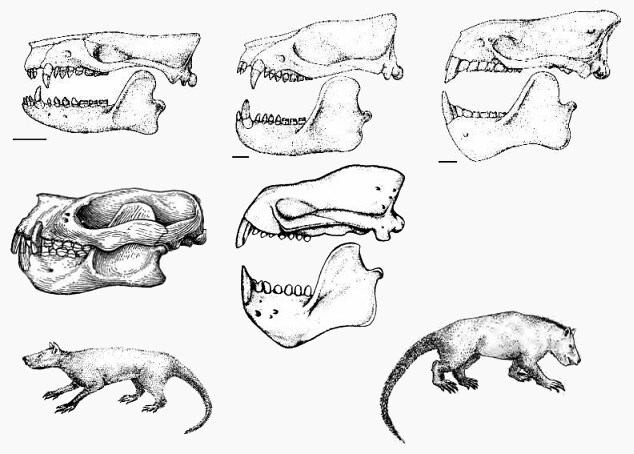
Тениодонты. Сверху — черепа: верхний ряд (слева направо) — Onychodectes, Wortmania, Ectoganus;
средний ряд — Psittacotherium, Stylinodon;
снизу — Onychodectes, Stylinodon
На последовательном ряду палеоценовых тениодонтов Taeniodonta можно видеть, как из подобного насекомоядным и опоссумам существа — Onychodectes — через собакоподобного зверя — Wortmania — может развиться странное чудище вроде Psittacotherium, Ectoganus или Stylinodon размером с медведя. Наверное, среди палеоценовых зверей тениодонты имели максимальную скорость эволюции. При этом они никем не рассматривались как особо прогрессивные млекопитающие.
Тениодонты могут служить наглядным примером, как можно быстро специализироваться и потерять возможность стать "настоящим приматом".
Другой пример быстрейшей специализации — летучие мыши Chiroptera. Рукокрылые, вероятно, имелись уже в верхнем мелу Южной Америки и верхнем палеоцене Франции и Германии (Gingerich, 1987; Hand et al. 1994; Hooker, 1996), а однозначные представители нижнего эоцена мало отличимы от современных, причём обнаружены сразу в десятках видов на всех континентах, включая Австралию.

Происхождение рукокрылых
Замечательно, что зубы нижнеэоценовых летучих мышей почти идентичны зубам примитивных плацентарных типа Cimolestes и древнейших землеройковых, так что родство всех этих групп не представляет сомнения, что однозначно подтверждается данными генетики. Несмотря на то, что в генетико-кладистических схемах рукокрылые попадают в лавразиатериев Laurasiatheria, а приматы — в эуархантоглиресов Euarchontoglires, сходство двух этих групп всегда было очевидно всем систематикам, начиная с К. Линнея, и отразилось в создании группы "архонта" Archonta, объединяющей летучих мышей, приматов, тупайй и шерстокрылов. Сходство пращуров представителей "архонта" усиливалось древесным образом жизни предков летучих мышей и шерстокрылов и, как минимум, преадаптацией к нему у предков приматов и тупайй. Очевидно, потому и не удаётся выявить непосредственных нижнепалеоценовых или меловых предков летучих мышей, что их зубы неотличимы от зубов прочих примитивных зверей. Не исключено, что какие-то палеоценовые формы, известные лишь по зубам и считающиеся ныне приматами, плезиадаписовыми или какими-либо насекомоядными в широком смысле, при лучшей изученности окажутся примитивными летучими мышами. Пока у летучих мышей не было крыльев и эхолокации, мы считаем их "насекомоядными", когда же эти специализации появляются (судя по Onychonycteris finneyi, полёт возник раньше эхолокации (Simmons et al., 2008)), мы уже видим готовых рукокрылых.
Как и в случае с птицами и птерозаврами, машущий полёт летучих мышей возник очень быстро, и уловить момент его становления крайне трудно, для этого надо обладать невероятным везением.
Рукокрылые уникальны в том отношении, что первые этапы их эволюции отличались максимальными темпами, а последующие — крайне низкими (вернее, на уровне видо- и родообразования темпы были велики, но план строения уже на уровне семейств практически не изменился с нижнего эоцена); можно даже утверждать, что макроэволюция летучих мышей закончилась в то время, когда у приматов она только начиналась. Очевидно, что причиной этого стала адаптация к полёту. И без того небогатые заделы строения мозга первопредков были безнадёжно задавлены необходимостью облегчения веса; наглядно это выражается в быстром зарастании швов черепа, что было характерно уже для раннеэоценового Icaronycterys. О хватательной способности конечностей речь тоже не идёт, скорее уж о цеплятельной; нижнеэоценовый Onychonycteris имел когти на всех пальцах крыла, а остальные синхронные родственники — уже потеряли на двух или трёх.

Ископаемая летучая мышь Icaronycterys (ранний эоцен)
Правда, у рукокылых есть два существенных преимущества перед насекомоядными: они долго живут, а потому могут накопить богатый жизненный опыт, и очень общительны — вплоть до заботы о голодных сородичах у вампиров Desmodus rotundus. Но эти преимущества сводятся на нет малыми размерами мозга — дорогая плата за покорение небес. Удивительно, но за десятки миллионов лет ни одна летучая мышь не потеряла способности к полёту и не вернулась к наземному или древесному образу жизни (в фантастической фауне будущего изобретательный ум Д. Диксона нагрезил хищных наземных вампиров, ходящих на передних лапах и хватающих добычу задними, но этот инфернальный образ пока, к счастью, остаётся сугубо гипотетическим и остаётся на совести своего создателя).
Много шуму в своё время наделала так нываемая гипотеза "летающих приматов", согласно которой мегахироптеры Megachiroptera — крылановые — приобрели способность к полёту независимо от прочих летучих мышей — микрохироптеров Microchiroptera, да к тому же возникли из древнейших приматов (Pettigrew, 1986; Pettigrew et al., 1989; Pettigrew et al., 2008). В доказательство приводилось множество доводов, основными из которых был специфический тип нервного соединения сетчатки глаза с верхниками холмиками четверохолмия в среднем мозге — уникальный для приматов, шерстокрылов и крылановых, а также отсутствие эхолокации у подавляющего большинства последних, в отличие от мелких эхолоцирующих рукокрылых. Приводились и другие доказательства независимого возникновения макро- и микрохироптеров.
В определённый момент концепция "летающих приматов" уже почти взяла верх, но тут же потерпела сокрушительное поражение от генетиков, довольно убедительно доказавших монофилию рукокрылых (Mindell et al., 1991);
были сделаны попытки оспорить эти генетические результаты (Hutcheon et al., 1998), но они не приняты большинством систематиков. Впрочем, признание единого происхождения летучих мышей не может отвергнуть множество удивительных параллелей крылановых и приматов. Даже если эти сходства развились конвергентно, они слишком комплексны, чтобы быть совсем случайными; всё же эта ситуация — отражение крайней близости предков обоих отрядов. Нет ископаемых форм, которые бы "зависали" между рукокрылыми и приматами (описан африканский раннемиоценовый крылан Propotto leakeyi, чьё имя говорит само за себя (Simpson, 1967; Walker, 1967), но тут дело в путанице, а не промежуточности) — это следствие быстрой специализации первых.
Много рассуждений было потрачено на выяснение вопроса — были ли предки летучих мышей насекомоядными или фруктоядными. Зубы древнейших известных форм "насекомоядные", но палеоценовые вполне могли отличаться большей любовью к произведениям флоры. Незатихающие споры по этой проблеме, а также существование обоих видов питания среди современных рукокрылых — лишнее подтверждение зыбкости грани между двумя этими диетами, сколь бы различными они не казались.

Скелет ископаемой летучей мыши Onychonycteris (эоцен)
В целом, последовательность специализаций рукокрылых видится примерно такой: судя по примитивнейшей летучей мыши Onychonycteris, не имевшей развитой эхолокации (хотя есть и другое мнение, что у него могла быть "гортанная эхолокация" (Veselka et al., 2010)) и питавшейся насекомыми, эхолокация возникла позже полёта, а первой диетой были насекомые; другие синхронные рукокрылые тоже насекомоядные, но эхолоцирующие. Судя по отсутствию эхолокации у большинства фруктоядных крылановых и её наличию у некоторых представителей этой же группы (египетская лутучая собака Rousettus aegyptiacus эхолоцирует, щелкая языком), а также по её сохранению у фруктоядных и нектароядных микрохироптеров, эхолокация могла исчезать у фруктоядных форм, но не обязательно; эхолокация и насекомоядность есть у генетически близких к крылановым подковогубовых Hipposideridae, подковоносовых Rhinolophidae, ложновампировых Megadermatidae, свиноносовых Craseonycteridae и мышехвостых Rhinopomatidae; с другой стороны, насекомоядные неоднократно и независимо переходили к фруктоядности. С другой стороны, все современные насекомоядные формы имеют развитую эхолокацию. Судя по развитию усложнённой нервой связи сетчатки и четверохолмия именно у неэхолоцирующих крылановых и примитивному варианту у всех прочих рукокрылых, "приматный" вариант нервной системы возник у крылановых независимо. Все эти тонкости кажутся посторонними для проблемы происхождения приматов, но на самом деле имеют к ней прямое отношение.
Ведь общие предки подразумевают, что и приматы имели шансы развить схожие адаптации.
Хищные птицы против рукокрылых
Интересно также, что в качестве одного из лимитирующих факторов, сдерживавших раннюю эволюцию рукокрылых и загнавших их в ночной образ жизни, называется гнёт со стороны дневных хищных птиц (Rydell et Speakman, 1995; Speakman, 2001; Simmons et al., 2008). Дескать, в конце мела птиц было много, и млекопитающие не имели шанса освоить воздух. Правда, ещё были птерозавры (из меловых нам известны в основном морские рыбоядные, но имелись и другие, которые могли мешать жить птицам) и хищные динозавры, отчего не было мелких насекомоядных птиц. Звери, будучи интеллектуальными созданиями, вероятно, меньше мелькали и лучше прятались в лесной подстилке и ветвях деревьев, почему, собственно, успешно заняли насекомоядную нишу.

Энанциорнис (поздний мел)
Птицы серьёзно пострадали во время позднемелового катаклизма, бóльшая часть их линий безвозвратно исчезла — например, энанциорнисы Enantiornithes, доминировавшие в мезозое.
В раннем палеоцене птиц осталось очень мало, и у зверей появился шанс на полёт. Тут-то и возникли летучие мыши и шерстокрылы.
К концу палеоцена птицы оклемались, отрастили крючковатые клювы и загнали распоясавшихся зверей в ночь.
Более того, в конце палеоцена уже появляются совы — Ogygoptynx wetmorei из США и Berruornis orbisantiqui из Франции — хищные птицы стали преследовать зверей и ночью. В позднепалеоценовых слоях Китая найдены копролиты или погадки птиц с останками млекопитающих — наглядная иллюстрация описываемых событий. Если всё действительно было так, может, приматы просто не успели взлететь?
Летучие мыши вовремя подсуетились, а приматы безнадёжно отстали, оставив в современном человеке вечную тоску по свободному полёту?
Правда, вопрос этот тёмный, и каждый шаг изложенных рассуждений подвергается бурным обсуждениям.
Птицы в палеоцене имели потенциальный шанс на "мировое господство". Млекопитающие были малы и слабы, а птицы были столь же теплокровны и умны. Однако, летающие птицы не могут иметь большой мозг — летать мешает, — а наземные формы возникают обычно в условиях отсутствия хищников, что расслабляет и приводит к ещё большему упрощению мозга. Все современные нелетающие птицы, мягко говоря, не блещут интеллектом: страусы, киви, дронты и прочие подобные создания являются чуть ли не образцом неразумности. Кстати, большой вопрос — что мешает цефализации страусов Struthioniformes, Rheiformes и Casuariiformes? Наземность в открытой местности, всеядная диета, зачатки социальности — некий задел имеется, но, может, рост слишком большой?
Возможно, главными кандидатами на чуть больший уровень цефализации могли бы быть пингвины Sphenisciformes:
сложное плавание в воде, сопоставимое с полётом, требует развития мозга, но потенциально позволяет нарастить его массу. Однако, антарктическая среда, бедная на раздражители, не способствует усложнению поведения пингвинов, а специализация крыльев как ласт не оставляет никаких надежд на развитие трудовой деятельности.

Ископаемая хищная птица Paraphysornis (миоцен, Бразилия)
Ископаемые наземные птицы Азии, Европы, обеих Америк и даже Антарктиды — фороракосовые Phorusrhacidae и гасторнисовые Gastornithidae (они же диатримовые Diatrymidae) — становились успешными хищниками в отсутствие крупных бегающих хищников-млекопитающих. Гасторнисовые возникли в палеоцене и вымерли в эоцене, два этих долгих периода они терроризировали зверей Северной Америки, Европы и Китая. В Южной Америке и Антарктиде фороракосовые появились в среднем палеоцене и оставались главными плотоядными вплоть до появления тут плацентарных — кошачьих и псовых, заселившихся с севера в позднем плиоцене; тогда же фороракосы даже освоили Северную Америку, хотя и ненадолго; вымерли же они лишь в начале плейстоцена. Эти ужасные создания в эоцене жили и на территории современного Алжира, а в раннем олигоцене — современной Франции. В Австралии вплоть до плейстоцена аналогичную экологическую нишу занимали дроморнисовые Dromornithidae — сумчатые так и не смогли составить им адекватную конкуренцию, и это при том, что появились дроморнисовые только в олигоцене. Потенциально фороракосовые и гасторнисовые могли стать интеллектуальнее прочих птиц: хищный образ жизни предполагает некоторое напряжение мозгов, а наземность обеспечивает возможности их роста.
Но, видимо, полуметровый клюв перевешивал. Принцип "сила есть — ума не надо" воплотился в ужасных птицах так же явно, как и в их родственниках — хищных динозаврах.
Шерстокрылы — почти приматы…
Раз уж речь зашла о полёте, невозможно пройти мимо шерстокрылов Dermoptera — ближайших родственников приматов по всем возможным показателям. Близость этих групп настолько велика, что многие систематики склонны определять шерстокрылов как подотряд приматов, называя их "летающие лемуры"; генетически шерстокрылы ближе к приматам, чем тупайи. Однако, морфологически шерстокрылы близки и к насекомоядным — настолько, что некоторые авторы включали их в этот отряд (например: VanValen, 1967).

Ignacius clarkforkensis (поздний палеоцен, Северная Америка)
Раз уж речь зашла о полёте, невозможно пройти мимо шерстокрылов Dermoptera — ближайших родственников приматов по всем возможным показателям. Близость этих групп настолько велика, что многие систематики склонны определять шерстокрылов как подотряд приматов, называя их "летающие лемуры"; генетически шерстокрылы ближе к приматам, чем тупайи. Однако, морфологически шерстокрылы близки и к насекомоядным — настолько, что некоторые авторы включали их в этот отряд (например: Van Valen, 1967). Собственно, древнейшие шерстокрыловые, древнейшие приматы и древнейшие насекомоядные различимы столь плохо, что статус целого ряда семейств и подсемейств остаётся под вопросом: это Mixodectidae, Placentidentidae, Ekgmowechashalinae, Thylacaelurinae и Paromomyidae. Тонкости строения основания черепа, включая слуховую капсулу и особенности расположения сосудов и нервов, размеры, пропорции, выпрямленность и строение суставных поверхностей фаланг кистей и стоп, гребешки для прикрепления мышц на них, форма когтей у Ignacius и Phenacolemur интерпретировалось по-разному (Bloch et Silcox, 2001; Hamrick et al., 1999; Krause, 1991).
Строго говоря, доказательств отнесения этих зверюшек к плезиадаписовым или шерстокрыловым примерно одинаковое количество.
Всё же большинство современных палеонтологов считают паромомиид плезиадапиформами, а прочие упомянутые семейства — шерстокрылами.

Малайский шерстокрыл.
Современные шерстокрылы крайне специализированы по множеству признаков. В первую очередь, конечно, стоит упомянуть летательную перепонку, тянущуюся от щёк и ушей до кончика хвоста и занимающую промежутки между длинными пальцами на руках и ногах, благодаря которой шерстокрылы могут планировать на большое расстояние. Во-вторых, шерстокрылы имеют крайне специфическую зубную систему с редукцией на верхней челюсти первых резцов и клыкоподобностью вторых, а также очень оригинальным строением нижних резцов — в виде фестончатых гребешков, ориентированных горизонтально. Получается очень близкий функциональный аналог "зубной щётки" современных лемуров, у которых направленные вперёд нижние резцы и резцеподобные клыки используются для чистки шерсти (кстати, такая "щётка" — одно из главных отличий современных лемуроподобных от их предков адапиформов Adapiformes и одновременно — лучший довод в пользу монофилетичности лемуроподобных, исключая, возможно, руконожку; скелет же и прочие черты у лемуров и адапид почти идентичны, разве что мозг у лемуров побольше; вообще подобные "щётки" возникали у зверей неоднократно: уже упоминался кондиляртр Chriacus с таким же приспособлением).

Челюсть Plagiomene
Сейчас шерстокрылы представлены всего двумя видами, но c палеоцена по миоцен они были более разнообразны, причём почти все ископаемые формы известны из Европы и Северной Америки. Самые бесспорные шерстокрылы из них — плагиомениды Plagiomenidae. Ellesmene eureka была найдена на крайнем севере Канады, по соседству с Гренландией, в эоцене эта местность находилась на 76о cеверной широты — далеко за полярным кругом; хотя тогда там и было не в пример теплее, чем сейчас, и росли субтропические леса, полярной ночи всё же никто не отменял. Конечно, Ellesmene жила там не в одиночестве: в её фауне встречены и другие виды, похожие на шерстокрылов, паромомисы, грызуны, многобугорчатые, лептиктиды, пантодонты, тениодонты, креодонты, миациды, различные копытные, птицы, крокодилы, саламандры и прочие животные (West et Dawson, 1978). Все они освоили столь необычные условия и, судя по изобилию их останков, были весьма многочисленны.
Скелет плагиоменид до сих пор не найден, они известны почти исключительно по челюстям и зубам, так что не ясно, были ли они такими же ловкими планёрами, как и современные шерстокрылы. Череп Plagiomene в некоторых деталях заметно отличается от шерстокрылового, так что их родство не слишком тесное.

Volaticotherium (юра или ранний мел, Китай)
Удивительным образом Plagiomene по сложности слуховой капсулы и морфологии двураздельных нижних резцов схожи с прыгунчиками, по пневматизации основания черепа — с выхухолями Desmana moschata, по многобугорчатости моляров — с шерстокрылами, а по строению медиальной стенки слуховой капсулы — с приматами, в целом же строение оказывается неповторимым среди млекопитающих (MacPhee et al., 1989).
Специализации зубов могли возникнуть конвергентно в разных линиях, а вот строение основания черепа у Plagiomene совсем уникально и особенно непохоже на вариант шерстокрылов. Так что плагиомениды вполне могут быть очередной оригинальной "околоприматной" линией.
Идея планирующего полёта возникала много раз параллельно.
Древнейший пример среди млекопитающих — юрский или раннемеловой Volaticotherium antiquum из Китая, выделенный в самостоятельный отряд Volaticotheria (Meng et al., 2006). Столь древние звери, конечно, не имеют прямого отношения к приматам и шерстокрылам, они были малы и обладали примитивным мозгом. Полёт во всех своих вариантах ставил крест на возможности "приматизации".
Плезиадапиформы — предки приматов
Во многом схожи с шерстокрылами непосредственные предки приматов — плезиадапиформы Plesiadapiformes. Они, с одной стороны, имеют общий план строения приматов, с другой — отличаются рядом параметров. Например, плезиадапиформы имели когти на всех пальцах, большую мордочку и примитивный мозг, глазницы, ориентированные вбок, по крайней мере у части видов отсутствовало окостенение слуховой капсулы, многие обладали специализациями в зубной системе. Благодаря отпечаткам из французского местонахождения Менат известно, что плезиадаписы имели пушистый хвост — не слишком характерный для приматов. Посему включение их в приматов или исключение из этого отряда — вопрос совести, а не таксономии. В настоящее время их всё же чаще выделяют в самостоятельный отряд Plesiadapiformes, но близость к приматам подчёркивается объединением тех и других в рамках надотрядной группировки Primatomorpha.
Относительно типа передвижения и образа жизни плезиадапиформов велись споры. Одни считали их наземными (Gingerich, 1976, 1984), другие — древесными (Szalay et Delson, 1979), однако ж отсутствие срастаний костей, гибкость конечностей и пропорции кисти однозначно свидетельствуют в пользу второго варианта. Впрочем, древесность древесности рознь: плезиадапиформы не умели бегать и скакать по веткам так же резво, как делают это сейчас обезьяны. Может, потому они и приобретали специализации зубов, подобные грызуньим — при пониженной мобильности надо извлекать из доступных ресурсов максимум выгоды? Неспроста зубная система, аналогичная системе многобугорчатых — с резко выступающими резцами, крупным режущим премоляром и перетирающими или давящими молярами — возникала неоднократно среди примитивных приматоподобных существ: у саксонеллид Saxonellidae, карполестоидов Carpolestoidea и фенаколемуриновых Phenacolemurinae. Наблюдая, сколь неряшливо едят обезьяны — без конца роняя что-то, выедая лишь самые вкусные кусочки и оставляя массу объедков — над этим можно задуматься.
Обезьяны всегда могут быстро переместиться и найти новые вкусняшки, а медленные плезиадапиформы, грызуны и многобугорчатые вынуждены довольствоваться тем, что есть на месте и грызть до победы.
Зубная система специализированных плезиадапиформов не была идентична таковой грызунов. У плезиадапоидов Plesiadapoidea верхние резцы имели бугорки и не были самозатачивающимися. От ранних форм к поздним доля мягкой пищи — фруктов и особенно листьев — в рационе, вероятно, увеличивалась (Boyer et al., 2010a). Плезиадапоиды были одними из самых успешных животных своего времени, в некоторых местонахождениях они оказываются наиболее встречаемыми из млекопитающих; это отразилось и в распространении: в Европе, Северной Америке, Пакистане и Китае с нижнего палеоцена по нижний эоцен. Размеры их сильно варьировали — самые мелкие были как белка, а некоторые были довольно крупными — размером с крупную кошку. Не слишком родственны, но экологически схожи с плезиадапоидами микросиопиды Microsyopidae, жившие с верхнего палеоцена по средний эоцен в Северной Америке; среди них встречались животные размером с бобра.

Carpolestes simpsoni (палеоцен)
Карполестоиды Carpolestoidea почти полностью перешли на питание волокнистыми плодами, орехами и стеблями, благодаря чему приобрели плоские и высокие премоляры с пильчатым краем. У них имелись и продвинутые приматные черты: например, большой палец руки был снабжён ногтем, а не когтем, что подразумевает и бóльшую чувствительность; впрочем, и карполестоиды были медлительными животными и не могли быстро прыгать по ветвям. Они существовали не так уж долго — в пределах среднего и верхнего палеоцена Северной Америки, а последние представители доживали свой век в нижнеэоценовых лесах Китая и среднеоэценовых — Пакистана. Карполестоиды никогда не были многочисленны и исчезновение их может быть связано с климатическими колебаниями или конкуренцией с более крупными животными, поскольку карполестоиды были одними из самых небольших плезиадапиформов — размером с мышь или крысу.
Пикродонтиды Picrodontidae, с их морщинистыми многобугорковыми, очень широкими — особенно первыми — и совсем не стёртыми молярами, стали очень специализированными фруктоядами, а может, даже питались преимущественно нектаром, пыльцой и древесным соком. Череп Zanycteris резко сужался спереди, этим пикродонтиды были похожи на современных мышиных лемуров Cheirogaleinae и хоботноголовых кускусов Tarsipedidae, питающихся нектаром. Пикродонтиды, видимо, дальше всех из плезиадапиформов ушли от исходной насекомоядности. Забавно, что зубы пикродонтид столь необычайно похожи на зубы летучих мышей, что в 1935 году Дж. Симпсон даже предполагал, что пикродонтиды относятся к рукокрылым (Simpson, 1935).
Паромомиоиды Paromomyoidea имели квадратные уплощённые моляры, приспособленные для перетирания фруктов, впрочем, в диете немалую роль могли играть древесный сок и насекомые. В последовательном ряду палеоценовых родов наблюдается быстрое удлинение и истончение нижних резцов с появлением промежутка — диастемы — между резцами и премолярами. На основании строения изолированных костей конечностей предполагалось, что паромомииды имели летательную перепонку, как шерстокрылы, но обнаружение и изучение целых скелетов показало, что эти звери были больше похожи на белок, а не на летяг.
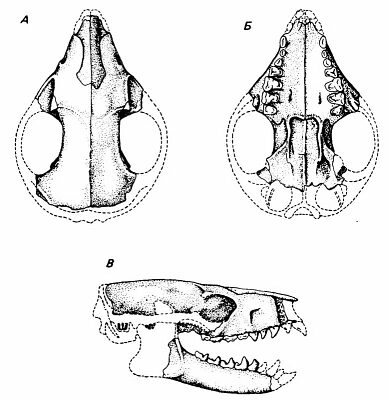
Череп паромомиоидового примата Palaechthon (средний палеоцен)
Все эти специализированные плезиадапиформы, конечно, не были предками приматов, но известны и более генерализованные их варианты, например, средне-верхнепалеоценовые североамериканские палеохтониды Palaechthonidae. У них сохранялась исходная для приматов зубная формула, не было диастемы между резцами и премолярами, отсутствовали гипертрофии каких-либо зубов. Строение черепа Palaechton nacimienti было самым примитивным среди всех плезиадапиформов: глазницы были маленькие, ориентированы вбок и широко расставлены, заглазничное сужение не выражено, обонятельные луковицы большие; судя по увеличенному подглазничному отверстию, имелись длинные вибриссы. Ориентировались палеохтониды в основном с помощью осязания, слуха и обоняния, а не зрения. Судя по всему, вели они в основном наземный образ жизни. В немалой степени все эти черты были связаны с малыми размерами тела и насекомоядностью палеохтонид (Kay et Cartmill, 1977). Показательно, что группа палеохтонид — вовсе не древнейшая среди плезиадапиформов; с одной стороны, она сохраняла исходную насекомоядность и экологически схожа с тупайями, с другой — по времени близка к настоящим приматам границы палеоцена и эоцена. Вряд ли настоящих приматов дали палеохтониды с их особенностями, но это должны были быть очень похожие на палеохтонид существа, только более древесные и всеядные.
Насекомоядность вполне сочеталась с древесностью у микромомиид Micromomyidae, которые были самыми мелкими плезиадапиформами — намного меньше современных мышиных лемуров. При их размере другая диета, нежели насекомые, млекопитающим резко противопоказана. Эти крошечные зверюшки жили в конце палеоцена и начале эоцена в Северной Америке.
Позднейшей группой насекомоядных плезиадапиформов были толиапиниды Toliapinidae (Hooker et al., 1999). Они появились лишь в конце палеоцена и дожили до среднего эоцена. Большинство видов, включая примитивнейшего Sarnacius gingerichi, найдено в Европе, но уже в начале палеоцена Altiatlasius koulchii мигрировал в Марокко, а самый поздний — Seia shahi — обнаружен в Пакистане. Altiatlasius koulchii первоначально был описан как древнейший настоящий примат семейства Omomyidae, но позже пересмотрен как плезиадапиформ. Толиапиниды были весьма схожи с микромомиидами по образу жизни и являются их полным "старосветским" аналогом; к сожалению, от тех и других найдено очень немного останков.
Плезиадапиформы имели мозг гораздо меньший, чем у приматов таких же размеров, но бóльший, чем у других зверей. Возможно, важнейшим признаком, обеспечившим в далёком будущем и наше появление, стало соотношение жевательной мускулатуры и мозговой коробки. Жевательные мышцы крепятся у млекопитающих на сагиттальном гребне, тянущемся по верху мозговой коробки; таким образом, они окружают её, уже с младых ногтей своим давлением способствуют раннему зарастанию швов черепа и препятствуют росту мозга. У приматомофов, начиная с плезиадапиформов, стала усиливаться тенденция к ослаблению обоняния и развитию стереоскопичности зрения. Это привело к повороту глазниц вперёд, уменьшению обонятельных луковиц и решётчатой кости, а в итоге — появлению заглазничного сужения черепа в том месте, где у всех приличных зверей находится обонятельный мозг (для гоминид сильное сужение — примитивный признак, а для приматов как отряда — прогрессивный, таковы причуды эволюции).
Таким образом, значительная часть сагиттального гребня оказалась впереди от мозговой коробки, а коробка, соответственно, сдвинулась от мышц назад, освободившись от его гнёта и получив возможность увеличиваться более долгий срок в онтогенезе. Фактически, редукция мозга привела к росту мозга!
Парадокс, но факт.
Впрочем, прогрессивные особенности плезиадапиформов не уберегли их от вымирания. Традиционно и по-умолчанию считалось, что в их исчезновении повинны более продвинутые настоящие приматы. Однако изучение частот встречаемости останков рисует другую картину (Fleagle, 1999). Резкий спад численности плезиадапиформов начинается совершенно синхронно со столь же резким подъёмом численности первых грызунов, тогда как лишь через некоторое время появляются и начинают распространяться полуобезьяны. Стало быть, плезиадапиформы проиграли эволюционную гонку грызунам, зато своим исчезновением освободили дорогу полуобезьянам. Таким образом, отличия плезиадапиформов от приматов оказываются крайне существенными: то, чего не было у первых, не позволило им успешно конкурировать с грызунами; то же, что приобрели вторые, вывело их на новый уровень, недоступный грызунам.
Среди плезиадапиформов особняком стоит семейство Adapisoriculidae. Особенности его представителей позволяли разным исследователям включать его в сумчатых, насекомоядных, лептиктид или миксодектид, а также считать предковым для тупайй. Адаписорикулиды назывались и как возможные предки афросорицид Afrosoricida, включающих современных златокротов Chrysochloridae и тенреков Tenrecidae (Seiffert, 2010). Несмотря на существенные различия с плезиадаписовыми в зубной системе, строение посткраниального скелета у плезиадапиформов, шерстокрыловых и адаписорикулид весьма схоже (Smith et al., 2010); судя по всему, это были древесные насекомоядные животные. Более того, плечевая кость адаписорикулид имеет промежуточное строение между вариантами меловых "кондиляртр" и кайнозойских эуархонт, причём в ряду самих адаписорикулид обнаруживается постепенный переход от более примитивного варианта мелового Deccanolestes к более эуархонтовому у палеогеновых форм (Boyer et al., 2010b). Если считать адаписорикулид приматами или приматоморфами, а Deccanolestes hislopi считать адаписорикулидом, то эта группа становится самой древней среди приматоморфов, поскольку указанный вид обнаружен в позднемеловых отложениях Индии.
Учитывая, что Индия в то время была островом, удалённым от всех других земель, а Purgatorius в то же время жил в Северной Америке, пути миграций становятся совсем непонятными.
Замечательно, что адаписорикулиды найдены в нижнем и верхнем палеоцене и нижнем эоцене Европы, а также верхнем палеоцене и нижнем эоцене Северной Африки, но ни малейших их следов нет в хорошо изученных фаунах Северной Америки и главной части Азии. Таким образом, либо адаписорикулиды были до крайности приматоподобными сумчатыми или насекомоядными, либо приматы могли возникнуть не в Северной Америке. Сами адаписорикулиды могли возникнуть в разных местах: древнейшие представители найдены в верхнем мелу Индии (несколько видов одного или двух родов, отсюда они могли попасть в Европу через Восточную Африку или острова моря Тетис), но самое большое разнообразие обнаруживается в нижнем палеоцене Европы (явно они появились тут раньше, но меловые европейские млекопитающие известны плохо, так что в Индию и Африку могли попасть отсюда), а примитивнейший вид Afrodon chleuhi известен из верхнего палеоцена Марокко (хотя более ранних в Африке пока не нашли, потенциально они могли распространиться отсюда и в Индию, и в Европу) (Smith et al., 2010; De Bast et al., 2012). Не исключено, что предки адаписорикулид жили в Европе и оттуда попали в Северную Америку, только там стали пургаториусами и, позже, плезиадапиформами. С другой стороны, адаписорикулиды могут быть предками современных шерстокрылов; из Индии они могли попасть в Юго-Восточную Азию, когда эти две части суши соединились (Smith et al., 2010). С другой стороны, даже отнесение рода Deccanolestes к плацентарным вызывает сомнения; согласно некоторым кладистическим расчётам, он вместе с прочими адаписорикулидами может представлять очень архаичную линию (намного примитивнее, чем Cimolestes, Zalambdalestes и Purgatorius), восходящую к корням всех эутериев, поздние представители которой сохранили примитивные черты до весьма поздних времён (Goswami et al., 2011). Впрочем, адекватность результатов кластерного анализа, применявшегося в подобных исследованиях по выявлению филогении, мягко говоря, спорна, что можно видеть, например, при сравнении выводов двух групп палеонтологов: в одной работе адаписорикулиды оказались предками тенреков (Seiffert, 2010), во второй — ответвлением древнейших и примитивнейших эутериев, даже близко не родственных ни тенрекам, ни эуархонтам (Goswami et al., 2011). Как бы то ни было, существование адаписорикулид свидетельствует, во-первых, о возможности межконтинентальных обменов фаун в позднем мелу и раннем палеогене, а во-вторых, о существовании примитивных групп, близких к предкам приматов, на осколках Гондваны — в Африке и Индии.
Приматоморфы
Приматоморфы — одна из самых древних или даже самая древняя группа млекопитающих из доживших до современности. Единственный моляр Purgatorius ceratops из верхнемелового слоя Монтаны мог быть внесён сюда из более позднего нижнепалеоценового (Lofgren, 1995), но в любом случае в раннем палеоцене приматоморфы уже существовали и бурно эволюционировали. Уже тогда среди плезиадапиформов мы видим массу специализированных форм.

Purgatorius — древнейшее приматоподобное млекопитающее (верхний мел — ранний палеоцен, США)
Быстрая специализация сопровождала появление каждой мало-мальски крупной группы приматов, настолько, что даже берёт удивление: от кого же возникали последующие группы, кто и где доносил до новых времён генеральный план строения (кстати, эта фраза — идеальный кандидат для цитирования креационистами, которые забудут процитировать следующее предложение)? Однако ж, существование доныне тупайй ставит всё на свои места: коли этот пресловутый "генеральный план" и сейчас скачет по веткам в лесу, для эволюции приматов не всё ещё потеряно. Речь, конечно, о тупайях.
Тупайи Scandentia в своём облике удивительно точно сохранили признаки фактически позднемеловых примитивных плацентарных.
Неспроста они регулярно оказывались в составе насекомоядных (например: Van Valen, 1967). Эти животные всеядны с упором на насекомоядность; преимущественно древесны, но некоторые наземны, хотя и они прекрасно лазают по деревьям; почти все активны днём. На пальцах у них когти, лапки не хватательные и большой палец не противопоставлен прочим, глаза находятся по бокам головы, обоняние отличное, мозг примитивный. Однако ж ведущие органы чувств — зрение и слух; на пальцах имеются дактилоскопические узоры, хоть и примитивные, зубы похожи на зубы лемуров и имеется такой специфический для лемуров орган как подъязык — вырост под языком с зазубренным краем, используемый для чистки передних нижних зубов. Тупайи наглядно демонстрируют нам примитивность приматов — эти два отряда весьма схожи. Древнейшие настоящие тупайи — Eodendrogale parvum — известны из среднего эоцена Китая; возможно, немного древнее неопределённые и вообще сомнительные тупайи из Турции (которые, строго говоря, могут быть кем угодно от сумчатых до копытных). Палеоценовые формы нам неизвестны, но примитивный план строения современных представителей не оставляет сомнения, что эти зверьки возникли намного раньше эоцена и должны восходить к меловым насекомоядноподобным млекопитающим.

Череп и нижняя челюсть Anagale
В качестве древнейших тупайй назывались анагалиды (конкретно — отряд Anagalida, а не когорта с таким же названием, но с гораздо бóльшим содержанием; чтобы избежать путаницы, отряд иногда называют Anagaliformes или Anagaloidea), но масса их специализаций в строении нижней челюсти, зубов и конечностей — включая раздвоенные когти на передних и копытца на задних ногах — позволяют говорить, что эта группа весьма отдалённо родственна "архонтам". Скорее всего, анагалиды возникли не от животных, похожих на Cimolestes, как "архонты", а от зверей типа Zalambdalestes (которые по этой причине иногда включаются в состав отряда анагалид). Даже экологически анагалиды отличались от предков приматов и прочих подобных групп; об этом свидетельствуют странная форма зубов и специализация конечностей.

Череп пиротерия Pyrotherium sorondoi (олигоцен, Южная Америка)
Впрочем, все эти особенности не мешают многим учёным считать анагалид предками грызунов и зайцеообразных;
они же могли быть предками огромных южноамериканских ксенунгулят Xenungulata и слоноподобных пиротериев Pyrotheria, что показывает великие возможности эволюции (Отдельный вопрос — как азиатские анагалиды оказались в Южной Америке? Поэтому некоторые палеонтологи считают предками ксенунгулят и пиротериев кондиляртр). Учитывая, что грызуны с зайцеообразными возникли чуть позже, чем "архонты" разделились на тупайй, шерстокрылов и приматов, но из того же филогенетического ствола, анагалид можно рассматривать как потомков тех же древних "архонт". Не зря некоторые систематики включают в когорту или надотряд Anagalida прыгунчиков, грызунов и зайцеобразных и объединяют их с Euarchonta (в отличие от Archonta, сюда не входят рукокрылые, но входят тупайи, шерстокрылы и приматы) в более крупный таксон Unguiculata. В некоторых системах прыгунчики оказываются в стороне, тогда Anagalida называется Glires, Euarchonta — Primatomorpha, а Unguiculata — Euarchontoglires. Однако ж, всё это — спор о словах, сути филогении это не меняет. Палеоценовые анагалиды жили в Азии и там могли в некоторой степени заменять приматов, хотя уже говорилось, что их экологические отличия достаточно очевидны. Вероятно, некоторые анагалиды выкапывали еду из земли, хотя в пределах отряда имеется достаточное разнообразие.
Прыгунчики
Прыгунчики Macroscelidea — особая тема. Эти небольшие зверушки традиционно относились к насекомоядным, на основании наличия слепой кишки они сводились с тупайями в группу Menotyphla, по строению предплюсны объединялись с зайцеобразными, по строению черепа, зубов и по эмбриологическим данным включались в анагалид, палеонтологические данные роднят прыгунчиков с палеоценово-эоценовыми североамериканскими кондиляртрами семейств Apheliscidae или, менее вероятно, Hyopsodontidae (и даже конкретнее — подсемейства Louisininae).

Короткоухий слоновый прыгунчик
Генетические данные — теоретически самые точные — выносят их в афротериев — группу африканского происхождения. Одно из наглядных отличий строения прыгунчиков от насекомоядных — большие размеры глаз, а поведения — дневной образ жизни; сходство с тупайями вообще довольно отдалённое. Показательно, что наиболее родственные прыгунчикам кондиляртры семейства Apheliscidae — Apheliscus и Haplomylus — одновременно весьма напоминают ежиных рода Macrocranion (Penkrot et al., 2008), а строение древнейших эоценовых прыгунчиков позволило предположить их близкое родство с Hyopsodontidae (Simons et al., 1991; Tabuce et al., 2001), тогда как его представители Hyopsodus и Sarcolemur первоначально были описаны как примитивные приматы. Посему совсем не странно, что, несмотря на генетическое расхождение, масса особенностей прыгунчиков сближает их с приматами. Важным представляется сравнительно сильное развитие головного мозга и относительно сложное поведение.

Macroscelides proboscideus (короткоухий прыгунчик)
Однако ж, существенно, что прыгунчики рано специализировались к прыганию по земле, отчего берцовые кости срослись, а большие пальцы и на кистях, и на стопах редуцировались; кости предплечья прижаты друг к другу, отчего способности к пронации-супинации резко ограничены.
Понятно, что хватательная функция кистей в итоге почти отсутствует.
Прыгунчики с самого своего возникновения и доныне питались преимущественно насекомыми, хотя некоторая доля растительной пищи в рационе тоже имеется; наличие слепой кишки, в отличие от ежей и землероек, показывает, что значение растительной пищи в прошлом могло быть бóльшим, чем сейчас. Миоценовый род Myohyrax имел зубы, подобные зубам грызунов и приспособленные к перетиранию растительной пищи, так что прыгунчики принципиально не были заперты в эволюционном тупике специализации. Такая смешанная диета вполне вероятна для древнейших приматов, но наши предки забрались на деревья, а прыгунчики освоили землю.
Невероятные тенреки
Впрочем, природа богата на выдумки. Древесные родственники у прыгунчиков тоже имеются — среди тенреков Tenrecidae. По ряду черт тенреки крайне архаичны; например, у них имеется клоака, но нет мошонки, а обмен веществ очень низкий, так что они почти холоднокровны.

Тенрек Hemicentetes semispinosus
Хватает и черт специализации: у тенреков нет скуловой кости, ежовые и полосатые тенреки Echinops telfairi, Setifer setosus и Hemicentetes semispinosus обзавелись иглами наподобие ежей, у некоторых слюна, вероятно, ядовита; в засушливый сезон многие виды впадают в спячку. Тенрековые вообще весьма разнообразны: среди них есть аналоги опоссумов и ежей, землероек и кротов, выхухолей и выдр. Питание у всех преимущественно насекомоядное, хотя при случае едят плоды; образ жизни ночной. Древесные формы принципом своей организации напоминают предков приматов. Любопытно, что тенреки миллионы лет сосуществуют на Мадагаскаре и в Африке с лемуровыми. Однако ж, тенреки и лемуры не конкурируют напрямую и не вытесняют друг друга. Тенреки в среднем мельче и более насекомоядны, а главное — у них куда более многочисленное потомство.
Хотя есть виды, у которых бывает 1–4 детёныша, у большинства их — около десятка, а рекорд принадлежит обыкновенному тенреку, рожающему до 32 малюток.

Обыкновенный тенрек Tenrec ecaudatus
Не случайно и количество сосков у этого вида — 29 — рекордно для млекопитающих. Понятно, что при максимальных длине и весе около 40 см и 2,5 кг (обычно же — в полтора раза меньше) размер детёнышей крайне невелик, есть проблемы и с качеством воспитания. При продолжительности жизни максимум несколько лет — в неволе до полутора десятков лет — возможности нарастить сколь-либо значимый мозг и обзавестись богатым жизненным опытом стремятся к нулю. Все эти показатели крайне контрастируют с лемурами, у большинства из которых обычно рождается один детёныш с максимумом у карликовых и мышиных лемуров, которые могут иметь до четырёх (и у них чаще рождается двойня). При этом лемуры в среднем заметно крупнее тенреков, а продолжительность жизни — больше 20 или даже 30 лет — гораздо внушительнее, отчего размеры мозга велики уже при рождении и заметно увеличиваются после. Как итог — социальное поведение тенреков и лемуров несопоставимо. Некоторые виды тенреков социальны, но выражается это лишь в том, что для сна несколько зверьков сцепляются в один шар, но кормятся они всё же по-одиночке. Не все лемуры общительны, но некоторые виды демонстрируют такой уровень социальности, какой тенрекам и не снился.
Афротерии
Группа афротериев Afrotheria вообще представляет уникальный пример возникновения крайне разнородных групп из одного корня. Корень этот разветвился в Африке примерно тогда же, когда в Евро-Америке возникли эуархонты-приматоморфы или даже конкретно приматы, однако последние остались гораздо однообразнее. Афротерии же дали наземных насекомоядных прыгунчиков Macroscelidea, подземных златокротов Chrysochloridae, роющих, наземных, древесных и полуводных тенреков Tenrecidae, трубкозубов Tubulidentata, даманов Hyracoidea, хоботных Proboscidea и сугубо водных сирен Sirenia.

Афротерии: 1. Трубкозуб 2. Дюгонь 3. Слоновый прыгунчик 4. Ламантин 5. Златокрот 6. Даман 7. Слон 8. Тенрек.
Уже по такому соотношению видно, что афротерии развивались в первоначальной изоляции от других групп млекопитающих. Видимо, в меловом периоде в Африке сложилась ситуация, аналогичная более поздней "великолепной изоляции" южноамериканской фауны. Приматоморфы же жили в окружении прочих бореоэвтериев, которые не давали им занять новые экологические ниши. Приматы очень рано — уже в среднем палеоцене, 57–60 млн.л.н. — появились в Африке; характерно, что древнейшая находка сделана в Марокко (Altiatlasius koulchii). Видимо, к этому времени афротерии были уже достаточно разнообразны и развиты, чтобы не проиграть в конкурентной борьбе бореоэвтериям, но аналогов приматов среди афротериев ещё не было.
История повторилась 26–27 млн.л.н. в Южной Америке — предыдущих миллионов лет не хватило, чтобы из опоссумов или грызунов возникло что-то подобное приматам.
Впрочем, в палеоцене разница афротериев и бореоэвтериев была слишком незначительной. И древнейшие прыгунчики (Chambius kasserinensis из нижнего-среднего эоцена Туниса, Nementchatherium senarhense из среднего-позднего эоцена Алжира, Herodotius pattersoni из позднего эоцена Египта), и древнейшие слоны (Eritherium azzouzorum из начала позднего палеоцена — 60 млн.л.н., Phosphatherium escuillei с границы палеоцена и эоцена — 56 млн.л.н., оба из Марокко, а также многочисленные роды и виды из эоцена Алжира, Египта и Ливии) обнаруживают явное родство с североамериканскими кондиляртрами (Hartenberger, 1986; Simons et al., 1991; Gheerbrant et al., 1996; Tabuce et al., 2001; Penkrot et al., 2008; Gheerbrant, 2009 и др.), причём хотя все они известны и из Африки, но из самой северной её части. Древнейший неназванный даман известен с границы палеоцена и эоцена Марокко, Seggeurius amourensis жил в нижнем эоцене Алжира (Court et Mahboubi, 1993; Barrow et al., 2010), более поздние эоценовые роды тоже североафриканские — из Марокко, Алжира, Туниса, Египта; их строение не свидетельствует очевидным образом о родстве с кондиляртрами, хотя таковое часто постулируется. Древнейшие тенрекоподобные животные (Todralestes variablis из позднего палеоцена Марокко, Dilambdogale gheerbranti из позднего эоцена Египта, а также более поздние Widanelfarasia и Qatranilestes оттуда же) тоже известны из Северной Африки, хотя ближе к адаписорикулидам Индии, Северной Африки и Европы, нежели кондиляртрам (Seiffert, 2010). Трубкозубы — самые специализированные из афротериев — не обнаруживают с прочими представителями этой группы какого-либо морфологического сходства, а происхождение их крайне туманно, поскольку все ископаемые формы очень поздние и очень похожи на современного трубкозуба; при всём этом, происхождение трубкозубых от кондиляртр не вызывает у палеонтологов сомнений; совпадения в строении с панголинами могут быть отнесены на счёт конвергенции. Учитывая, что кондиляртры были широко распространены в Азии, Европе, Северной и даже Южной Америке, всё это может свидетельствовать о евро-американском либо азиатском происхождении афротериев или вообще полифилии этой группы. Близкие к предкам слонов кондиляртры жили в Азии: например, фенаколофиды Phenacolophidae, из которых древнейшая — Minchenella grandis — известна из позднего палеоцена Китая, а чуть более поздний монгольский Phenacolophus родственен также эмбритоподам Embrithopoda, примитивнейшие представители которых найдены в Румынии и Турции, а в развитом виде — в Египте; антракобуниды Anthracobunidae, которые обычно относятся непосредственно к хоботным, появляются в раннем эоцене Пакистана (Wells et Gingerich, 1983); фенаколофиды и антракобуниды, правда, могут быть предками не хоботных, а десмостилий Desmostylia (известных только с северных побережий Тихого океана), но в любом случае родственны. Схожая с кондиляртрами примитивнейшая среднеэоценовая сирена Prorastomus с Ямайки и другие эоценовые сирены имели пятый премоляр, тогда как у всех кайнозойских млекопитающих он исчез, так что отхождение линии сирен от прочих афротериев надо бы относить к меловому периоду.
Значит ли вся совокупность этих данных, что часть кондиляртр в генетическом отношении была афротериями? Надо ли относить корень афротериев в меловой период или эоцен? И возникли ли они в Африке, ведь родственные группы обнаруживаются и в Индии, и в Азии, и в Европе, и в Северной Америке? Или кондиляртры в принципе сборная группа, объединяемая палеонтологами больше в силу недостатка сведений?
Как понимать генетическое родство афротериев, если их палеонтологические корни столь разлаписты?
В любом случае, очевидно близкое сходство и, вероятно, родство мелких всеядных наземных и лазящих млекопитающих даже в эоцене, не говоря уж у меловом периоде и палеоцене.
Происхождение афротериев от единого предка, отстаиваемое генетиками, ещё требует серьёзного палеонтологического подтверждения и обоснования. По другую сторону Атлантического океана "великолепная изоляция" сработала гораздо однозначнее. Конечно, речь о зверях Южной Америки.
Древние млекопитающие Южной Америки
Древние южноамериканские фауны характеризуются изобилием экзотических отрядов копытных Meridiungulata: нотоунгуляты Notoungulata, астрапотерии Astrapotheria, пиротерии Pyrotheria, ксенунгуляты Xenungulata и литоптерны Litopterna; иногда выделяются и другие. Удивительно, но среди великого разнообразия всех этих полуфантастических зверей так и не возникли аналоги приматов. Были псевдолошади, псевдоверблюды, псевдоносороги, псевдослоны, псевдодаманы и масса прочих существ, конвергентных со "старосветскими", но "псевдоприматы" нам неизвестны. С одной стороны, этот факт облегчил жизнь первым широконосым приматам, с другой — остаётся вопрос: почему?

Череп Нотоунгулята Notostylops
Южноамериканские кондиляртры быстро дали массу копытных отрядов, большей частью крупноразмерных, грызуны хорошо освоили нишу мелких и средних растительноядных, но при великих возможностях, предоставляемых пышной тропической флорой, почти не дали древесных форм (только древесных дикобразов Erethizontidae), тем более не дали фруктоядных и с быстрой локомоцией. Может, южноамериканские грызуны не могли преодолеть конкуренцию опоссумов? Сомнительно. О конкуренции со стороны ленивцев речь вообще не идёт. Вероятно, грызуны уже были заметно специализированы к моменту попадания в Южную Америку и не смогли эффективно перестроиться. Может быть, ранние этапы заселения плацентарными Южной Америки пришлись на время, когда там преобладали степные ландшафты, а к моменту значительного расширения лесов фауна уже сильно специализировалась к наземной жизни? Такое предположение подтверждается великим разнообразием крупных южноамериканских копытных, адаптированных к жизни в открытой местности.
Не исключено также, что мы просто не знаем этих древних древесных грызунов Южной Америки, как это часто бывает с лесными лазящими формами по чисто тафономическим причинам;
однако, приматы нам известны из нескольких местонахождений и неочевидно, почему там не могли сохраниться останки других древесных животных. В любом случае, уровень интеллектуального развития южноамериканских млекопитающих был явно ниже, чем у прибывших сюда в олигоцене обезьян.

Девятипоясный броненосец
Неполнозубые-ксенартры Xenarthra всегда имели крайне примитивный мозг. Однако, потенциально у них не раз возникали неплохие шансы для "приматизации". Древесный образ жизни в сочетании с питанием высококалорийной пищей — как раз подходящий вариант. Муравьеды Vermilingua — тамандуа и карликовый, равно как и некоторые панголины Pholidota приспособились к этому неопределённо давно, начало их специализации может восходить чуть ли не к меловому периоду. Учитывая великую роль питания термитами в становлении всеядности и трудовой деятельности австралопитеков и "ранних Homo" (Длусский, 1980; Backwell et d’Errico, 2001), можно было бы ожидать от муравьедов и панголинов некоего интеллектуального уровня, но увы! И те, и другие имеют ближайших родственников среди наземных роющих животных и имеют самое архаичное строение среди плацентарных: примитивную плаценту, очень простой мозг. Масса специализаций не позволяет надеяться на сколь-либо "разумное" будущее этих зверей. Тем более мало шансов у броненосцев Dasypodidae и ленивцев Bradypodidae — крайне специализированных зверей. Возможно, чуть больше шансов было у гигантских наземных ленивцев Megalonychidae, Megatheriidae и Mylodontidae, тем более что некоторые их группы демонстрировали удивительную экологическую пластичность, дав, в частности, плавающие морские формы Nothrotheriinae (Muizon et McDonald, 1995),
однако, гигантские размеры в сочетании с растительноядностью всегда заводили животных в эволюционный тупик.
Афротерии, неполнозубые-ксенартры и южноамериканские копытные в генетическом плане — отдалённая родня, иногда объединяемая в рамках группы атлантогенат Atlantogenata. Как было показано выше, во всех этих группах "приматизация" упорно не шла. Среди всего разнообразия атлантогенат лишь слоны могут похвастаться недюжинным интеллектом, но, скорее всего, это просто побочный эффект очень больших размеров, примерно как и в случае с китообразными. Огромные размеры слонов и их бесчисленные специализации — тумбообразные ноги, хобот, бивни, тяжёлые челюсти — не оставляют надежд на появление у них настоящего разума
Почему приматы? Предпосылки разумности…
Очевидно, что одной из важнейших черт приматов является их древесность. Однако, давно было замечено, что приматы обладают массой особенностей, отсутствующих у других древесных животных. Некоторые учёные предположили, что специфика приматов обусловлена ориентацией на зрение в совокупности с насекомоядностью и жизнью на ветках (Cartmill, 1972), но эта точка зрения подверглась основательной критике (Sussman, 1991). Уже самые первые приматоморфы имели склонность ко всеядности, чем они, собственно, и отличались от насекомоядных. Переход с преимущественно насекомоядной диеты на всеядную мог быть первым толчком к появлению приматов.
Редкие случаи насекомоядности приматов относятся как раз к сравнительно поздним временам: долгопяты являются одними из самых специализированных форм; насекомоядные приматы — галаго, потто, лори, долгопяты — ориентируются в основном на слух, а не на зрение. Слепая кишка имеется у всех приматов, свидетельствуя о их исходной всеядности. С другой стороны, специализированно растительноядные формы возникали среди приматов тоже не так уж часто.
Крайность в этом направлении представляют тонкотелые Colobinae, у которых в связи с исключительной листоядностью даже возникает многокамерность желудка, подобная варианту жвачных копытных Ruminantia;
но даже самые листоядные приматы стремятся получать более питательную пищу, регулярно поедая фрукты и насекомых; показано, что цветное зрение помогает современным колобусам выискивать среди зелени красноватые листья, более богатые белками, обезьяны предпочитают есть их, оставляя зелёные нетронутыми (Moffat, 2002). Возможно, так же вели себя и плезиадапиформы. Очевидно, именно поэтому ранние приматы — за исключением самых специализированных форм — не утеряли клыки, и мало кто из них обзавёлся большой диастемой между передними и задними зубами, столь характерной для сугубо растительноядных животных. Клыки сохранялись и по причине повышенной социальности приматов, так как играли важнейшую роль в регуляции внутригрупповых взаимоотношений.
Немалую роль сыграла редукция обоняния, неэффективного на деревьях, в сочетании с усилением роли зрения, осязания и, в меньшей степени, слуха. Жизнь в постоянном движении на качающихся кончиках веток, где приматы были в большей безопасности, требовала развития чувства равновесия, стереоскопичности зрения и развития структур мозга, ответственных за трёхмерное восприятие объектов и оценку расстояния. О взаимосвязи редукции обонятельных луковиц, стереоскопичности зрения, развития жевательной мускулатуры и роста мозга уже говорилось выше. Древнейшие приматы, видимо, были дневными животными, в связи с чем у них сохранилось цветное зрение — с его помощью в зелёной листве легче находить яркие фрукты и насекомых, нежели используя обоняние.
Жизнь на деревьях позволила также сохранить и развить хватательные способности кисти. Хотя некоторые приматы — потто, галаго, паукообразные обезьяны, игрунки, колобусы, гиббоны — в итоге приобрели крайне специализированные формы её строения, у большинства видов во все времена сохранялись умеренные пропорции и противопоставление большого пальца. Хватательная же функция кисти, в свою очередь, позволяет использовать её для манипуляции пищей, что опять же должно быть обеспечено развитием мозга.
Древесность бывает разная. Приматы в большинстве своём передвигаются по деревьям быстро, в основном прыжками, причём способны бегать по тонким ветвям и даже их кончикам (Orkin et Pontzer, 2011).
Резвые скачки в сложном мире качающихся веток должны иметь мощное обеспечение в виде большого мозга.
Конечно, на каждую стройную концепцию есть свои белки, не знающие о необходимости цефализации, но тенденция, тем не менее, очевидна. Секрет в том, что белки передвигаются в основном вдоль стволов и толстых веток; с дерева на дерево белки предпочитают перебираться или по почти соприкасающимся толстым ветвям или по земле, а длинные прыжки с тонких веток на другие тонкие ветки совершают крайне редко; даже летяги прыгают со ствола на ствол, большая прицельность при этом не нужна. Но и среди белок есть свои исключения. Восточная серая белка Sciurus carolinensis способна, в отличие от прочих белок, кормиться на кончиках тонких веток без адаптаций, свойственных приматам. Кроме того, есть тупайи, поссумы и хамелеоны, тоже кормящиеся на кончиках тонких веток, но не обладающие великим интеллектом, ногтями и бинокулярностью (хотя все они не прыгают подобно приматам).
Таким образом, жизнь на кончиках веток могла быть важнейшим фактором эволюции приматов только при условии отсутствия приспособлений к другим способам добывания пищи (Orkin et Pontzer, 2011)
— вывод замечательный, учитывая столь часто постулируемую экологическую и этологическую пластичность приматов!

Череп эоценового омомиида Tetonius
Собственно, все вышеперечисленные признаки и особенности являются главным препятствием для признания плезиадапиформов в качестве настоящих приматов. Ведь они обладали маленьким мозгом с хорошо развитыми обонятельными центрами; зрение, судя по направленным в стороны глазам, не было стереоскопичным; имелись когти на пальцах, их большой палец не противопоставлялся, а кисть была больше "цеплятельной", нежели "хватательной". Поэтому, несмотря на тот факт, что плезиадапиформы уже ушли от сугубой насекомоядности, а некоторые стали специализированными листо- и фруктоядами, многие систематики склонны считать первыми приматами только эоценовых адапиформов Adapiformes и омомиформов Omomyiformes.
Эволюция ранних приматов шла в тесной связи с эволюцией цветковых растений (Sussman, 1991). Даже стремление приматов забираться на кончики веток может быть связано с питанием цветами и плодами. В этой связи никак нельзя пройти мимо того факта, что главный "эволюционный бум" среди млекопитающих, согласно исследованиям по коннексии палеонтологических и генетических данных, случился отнюдь не в начале палеоцена, а намного раньше — около 80 миллионов лет назад (Bininda-Emonds et al., 2007; Meredith et al., 2011). Заметно меньший всплеск семействообразования приходится на время около 100 тысяч лет и примерно такой же — на начало палеоцена.
Тогда как последний очевидно связан с вымиранием динозавров, два древнейших примерно совпадают с появлением цветковых растений и установлением их доминирующей роли в мире растений.
Приматы по таким расчётам возникли от 90 до 70 — по самой новейшей разработке — миллионов лет назад. Что любопытно, согласно этой новейшей разработке приматы появились раньше тупайй, что крайне странно, учитывая очевидную примитивность последних (Meredith et al., 2011). Древнейшие — начиная со среднеюрской Juramaia sinensis и продолжая меловыми азиатскими и американскими видами — плацентарные или близкие к плацентарным звери в современной фауне больше всего схожи именно с тупайями, которые, таким образом, выглядят самыми примитивными плацентарными современности, по крайней мере в экологическом смысле (Luo et al., 2011). Даже насекомоядные экологически более прогрессивны, поскольку ведут преимущественно наземный образ жизни, тогда как исходным был, видимо, древесный. Хотя тупайй уже давно не включают в отряд приматов, всё же родство этих групп несомненно.
Палеоценовый мир был разделён на несколько больших географических областей. Южная Америка, Антарктида и Австралия представляли собой уже независимые части былой Гондваны, которым позже суждено было вновь соединиться. Северная Америка хотя и была отделена мелководным морем от Европы, всё же была объединена родственными фаунами (обмены фаун Европы и Северной Америки констатируются ещё в конце мелового периода (Martin et al., 2005)). Азия была отделена от Европы широким проливом и существовала сама по себе, хотя в последующем соединилась с Северной Америкой. Индия плавала посреди океана, а в эоцене воткнулась-таки в Азию. Африка, почти перерезанная вдоль глубоким заливом на месте Сахары, лежала недалеко от Европы, но напрямую с ней не сообщалась, а между Африкой и Азией тянулась цепь островов наподобие современной Индонезии.
В каждой из этих областей возникли свои фауны, в каждой из которых возникли во многом похожие формы.
Приматоморфы зародились в Северной Америке; в позднем палеоцене они известны в Европе сразу во множестве форм, а судя по тому, что в среднем палеоцене приматоморфы добрались уже и до Африки, в Европе в это время они тоже должны были быть. В Азии приматы, возможно, появились тоже в среднем палеоцене, но китайский Decoredon anhuiensis может быть как омомиидом, так и анагалидом, и, с очень большой вероятностью, кондиляртром (возможно даже — уже поминавшегося семейства Hyopsodontidae), а достоверные приматы в Китае отмечены в позднем палеоцене, хотя вначале они там крайне немногочисленны и почти идентичны североамериканским. На Индостанском острове приматоморфы появились как минимум в раннем эоцене, если не принимать в расчёт спорных адаписорикулид. Фаунистические связи работали в разных направлениях, и трудно разобрать, насколько реконструированная зоогеографическая схема соответствует действительности.
Кем приматы не стали?
В палеоцене в разных регионах имелись свои потенциальные "приматы": в Азии — анагалиды, в Африке — тенреки, в Южной Америке — опоссумы, в Австралии — кускусы. И лишь в Европе и Америке зародилась группа приматоморфов. Только на границе палеоцена и эоцена — с глобальным потеплением и распространением тропических лесов — возникают настоящие приматы адапиформы Adapiformesи омомиформы Omomyiformes, которые чрезвычайно быстро расселились по Азии и Африке и дали в последующем все более поздние группы. Приматы стали приматами в результате целого ряда конкретных обстоятельств.
Огромную роль сыграли первичные морфологические и этологические специализации и конкурентные взаимоотношения с другими млекопитающими и птицами. Не будь великого множества прочих животных — приматы так никогда бы и ступили на путь "приматизации" и никогда не увенчались бы разумным "венцом".

Руконожка ай-ай Daubentonia madagascariensis
Среди приматов можно найти примеры конвергенции с самыми разными животными. Руконожка ай-ай Daubentonia madagascariensis имеет постоянно растущие резцы, как у грызунов. Мадагаскарские палеопропитеки Palaeopropithecus и бабакотии Babakotia, вымершие уже в историческое время, пропорциями конечностей и способом локомоции мало отличались от ленивцев. Мартышки-гусары Erythrocebus patas и мезопитеки Mesopithecus скорее напоминают в этих отношениях газелей, а галаго Galago — тушканчиков. Зубы гигантопитеков Gigantopithecus на удивление похожи на зубы крупных травоядных, причём не только копытных, но и ископаемых гигантских панд. Мандрилы Mandrillus имеют впечатляющие клыки, по сути это — саблезубые обезьяны. Конвергенция касается не только строения, но и поведения. Так, толстохвостые лемуры Cheirogaleus впадают в спячку на сухой сезон, как делают и многие другие животные в неблагоприятные времена. Образ жизни и способ добывания пищи современных горилл Gorilla и, возможно, гигантопитеков Gigantopithecus, напоминают таковые халикотериев Chalicotheriidae, гигантских наземных ленивцев Megatherioidea и, в меньшей степени — нотоунгулятных хомалодотериев Homalodotheriidae Южной Америки. Так что у приматов было много адаптационных возможностей, которые они успешно использовали!
Однако, весьма поучительно знать не только экоформы, приобретённые приматами, но и те, которыми приматы так никогда и не стали.
Рассмотрение таковых показывает ограниченность экологической пластичности приматов и, одновременно, подчёркивает специфику этого отряда.

Юрское водное млекопитающее Castorocauda lutrasimilis
Среди всего разнообразия среди приматов ни разу не возникли специализированные водные формы. Носачи Nasalis larvatus, переходящие затопленные мангровые заросли по пояс в воде, макаки-крабоеды Macaca fascicularis, добывающие пищу в воде, и целебесские макаки Macaca nigra, плещущиеся в морском прибое, — даже не повод для сравнения с выхухолями, бобрами и капибарами. Приматы с самой свой зари были столь далеки от воды, что за десятки миллионов лет не смогли полюбить её всей душой. Человек в этом смысле — большое исключение, хотя и он стал человеком тогда, когда вышел осваивать горизонты саванн, а не морские просторы.
Многочисленные и модные домыслы о невероятной связи человеческих предков с водой, вплоть до постулирования необычайного родства с дельфинами, не имеют под собой особых оснований.
Древнейшие звери, приспособленные к водному образу жизни, известны уже из середины — Castorocauda lutrasimilis (Ji et al., 2006) — и конца юрского периода — Haldanodon exspectatus (Martin, 2005). Бесконечно далеки они от приматов!
Водные млекопитающие обычно довольно интеллектуальны (есть, конечно, дюгони Dugongidae и ламантины Trichechidae, но морская корова и есть корова, хотя и морская). В воде можно почти бесконечно наращивать размеры мозга, тем более что ресурсы позволяют. Китообразные Cetacea ещё в олигоцене достигли церебральных показателей — по абсолютным размерам и складчатости поверхности — уровня продвинутых гоминид (Lilly, 1977), а многие современные виды вдвое превосходят по ним человека. Однако, общеизвестной истиной является малая относительная величина мозга китообразных, а преобразование лап в ласты — не лучший зачин для освоения трудовой деятельности. Кроме того, ловля рыбы и креветок — не самый идеальный стимулятор умственной деятельности. Что бы энтузиасты и фантасты ни говорили о необычайных достижениях дельфинов, шансов на разумность у них немного. То же можно повторить и о ластоногих Phocidae, Otariidae и Odobenidae (или, обобщённо — и пусть генетики покосятся на меня! — Pinnipedia).
Не освоили приматы и роющий образ жизни, столь любимый ближайшими родственниками — насекомоядными и грызунами. Древнейшие специализированно-роющие звери известны уже из верхней юры — это Fruitafossor windscheffeli из Колорадо (Luo et al., 2005).
Современные шахтёры и спелеологи — первый шаг в столь новой для приматов области, как недра земли, но внутренние сомнения не дают поверить в скорое эволюционное обособление морлоков.
Приматы не стали хищными. Конечно, приматы возникли из насекомоядных существ и почти все не против съесть кого-нибудь не слишком опасного, но избыточной свирепостью обезьяны никогда не отличались, ведь и самые хищно настроенные долгопяты, лори и галаго постоянно едят фрукты. Человек — максимальное приближение к идеалу плотоядного, но даже светлый образ неандертальца-суперхищника в последнее время оказался замутнён фактом варки каш из ячменя и поедания фиников, бобов, кувшинок и сорго (Henry et al., 2011).
Приматы никогда не дали крупных наземных травоядных. Гориллы и особенно гигантопитеки, сколь бы велики и растительноядны они ни были, не могут сравниться с антилопами, носорогами и слонами; да и гориллы при всей своей тяжести не выходят из густых лесов и отлично лазают по деревьям. Приматы с самого начала своей эволюции не умели жить на земле. Показательно, что даже на Мадагаскаре с его бедной на хищников фауной лемуры не дали полноценных наземных видов. Лишь несколько приматов спустилось с деревьев, и лишь один — человек — успешно. Даже галаго, столь похожие на тушканчиков и живущие в буше, скачут в основном по ветвям, а не под ними. Этим приматы основательно отличаются от всех близких групп млекопитающих — грызунов, насекомоядных, афросорицид, ископаемых кондиляртр, лептиктид и многих прочих, включая даже тупайй. Лишь шерстокрылы и летучие мыши сопоставимы с приматами по степени "отрыва от почвы" и даже превзошли их в этом, но шерстокрылов всего два вида, а летучие мыши оторвались не только от почвы, но и от деревьев.
Неспециализированные?
Учитывая всё вышесказанное, нельзя согласиться с часто приводимым рассужением, что приматы — удивительно неспециализированные и генерализованные, что они "сделали ставку на специализацию к неспециализации", что они адаптируются силой мысли, а не морфологически. Среди мира млекопитающих приматы заняли вполне определённую экологическую нишу, которую ревниво оберегали весь кайнозой, крайне редко покушаясь на чужие вотчины. Приматы за свою историю дали массу удивительно специализированных форм; с другой стороны, многие древние и современные животные гораздо менее специализированы.
Секрет успеха приматов кроется в сочетании специализации к древолазанию и всеядности (с уклоном в растительноядность) с сохранением заметной доли экологической пластичности. Жизнь в трёхмерном мире на ветках деревьях привела к развитию зрительного и тактильного анализаторов с подавлением обонятельного, заставила напрягать мозги и пользоваться руками; одновременно не дала конечностям закостенеть и специализироваться по пути срастаний и редукций элементов. Разнообразное и достаточно калорийное питание способствовало загружению мозга нестандартными задачами и обеспечению энергетики для их решения. Не слишком малые размеры тела, малое число детёнышей и большая продолжительность жизни обеспечили возможности развития крупного мозга и наполнения его богатой иноформацией — жизненным опытом.
В комплексе с высоким уровнем социальности это дало приматам возможность развития интеллекта вплоть до человеческих высот и, хочется верить, бóльших.
Литература
Длусский Г.М. Некоторые соображения о начальных этапах антропогенеза // Вопросы Антропологии, 1980, вып. 66, стр. 85–90.
Кэрролл Р. Палеонтология и эволюция позвоночных. Т.2. М., Мир, 1993а, 283 с.
Кэрролл Р. Палеонтология и эволюция позвоночных. Ред.: В.И. Громова. Т.3. М., Мир, 1993б, 312 с.
Лопатин А.В. Строение черепа Archaeorycteseuryalis sp. nov. (Didymoconidae, Mammalia) из палеоцена Монголии и систематическое положение семейства // Палеонтологический журнал, 2001, № 3, сс.97-107.
Лопатин А.В. Особенности развития фауны мелких млекопитающих Азии в раннем палеогене // Экосистемные перестройки и эволюция биосферы. М., ПИН РАН, 2004, Вып.6, сс.87–96.
Основы палеонтологии. Т.13. М., Государственное научно-техническое издательство литературы по геологии и охране недр, 1962, 423 с.
Archibald J.D. et Averianov A. Phylogenetic analysis, taxonomic revision, and dental ontogeny of the Cretaceous Zhelestidae (Mammalia: Eutheria) // Zoological Journal of the Linnean Society, 2012, V.164, pp.361–426.
Backwell L.R. et d’Errico F. Evidence of termite foraging by Swartkransearly hominids // PNAS, 2001, V.98, № 4, pp.1358–1363.
Barrow E., Seiffert E.R. et Simons E.L. A primitive hyracoid (Mammalia, Paenungulata) from the early Priabonian (Late Eocene) of Egypt // Journal of Systematic Palaeontology, 2010, V.8, № 2, pp.213–244.
Bininda-Emonds O.R.P., Cardillo M., Jones K.E., MacPhee R.D.E., Beck R.M.D., Grenyer R., Price S.A., Vos R.A., Gittleman J.L. et Purvis A. The delayed rise of present-day mammals // Nature, 2007, V.446, pp.507–512.
Bloch J.I. et Silcox M.T. New basicrania of Paleocene-Eocene Ignacius: re-evaluation of the Plesiadapiform-Dermopteran link // AJPhA, 2001, V.116, pp.184–198.
Boyer D.M., Evans A.R. et Jernvall J. Evidence of Dietary Differentiation Among Late Paleocene — Early Eocene Plesiadapids (Mammalia,Primates) // AJPhA, 2010a, V.142, pp.194–210.
Boyer D.M., Prasad G.V.R., Krause D.W., Godinot M., Goswami A., Verma O. et Flynn J.J. New postcrania of Deccanolestes from the Late Cretaceous of India and their bearing on the evolutionary and biogeographic history of euarchontan mammals // Naturwissenschaften, 2010b, V.97, pp.365–377.
Cartmill M. Arboreal adaptations and the origin of the order Primates // The functional and evolutionary bioligy of primates. Ed.: R. Tuttleed. Chicago, Aldine, 1972, pp.97-122.
Court N. et Mahboubi M. Reassessment of Lower Eocene Seggeurius amourensis: aspects of primitive dental morphology in the mammalian order Hyracoidea // Journal of Paleontology, 1993, V.67, № 5, pp.889-893
De Bast E., Sige B. et Smith Th. Diversity of the adapisoriculid mammals from the early Palaeocene of Hainin, Belgium // Acta Palaeontologica Polonica, 2012, V.57, № 1, pp.35–52.
Gheerbrant E. Paleocene emergence of elephant relatives and the rapid radiation of African ungulates // PNAS, 2009, V.106, № 26, pp.10717-10721.
Gheerbrant E., Sudre J. et Cappetta H. A Palaeocene proboscidean from Morocco // Nature, 1996, V.383, pp.68–71.
Gingerich P.D. Cranial anatomy and evolution of early Tertiary Plesiadapidae (Mammalia, Primates). Univ.Michigan, Papers on Paleontol., 1976, V.15, pp.1-140.
Gingerich P.D. Primate evolution // Mammals. Notes for a Short Course. Eds.: P.D. Gingerich et C.E. Badgley. Univ.Tennessee, Studies Geol., 1984, V.8, pp.167–181.
Gingerich P. Early Eocene bats (Mammalia: Chiroptera) and other vertebrates in freshwater limestones of the Willywood formation, Clark’s Fork Basin, Wyoming // Contributions of the Museum of Paleontology at the University of Michigan, 1987, V.27, pp.275–320.
Goswami A., Prasad G.V.R., Upchurch P., Boyer D.M., Seiffert E.R., Verma O., Gheerbrant E. et Flynn J.J. A radiation of arboreal basal eutherian mammalsbeginning in the Late Cretaceous of India // PNAS, 2011, V.108, № 39, pp.16333-16338.
Hamrick M.W., Rosenma B.A. et Brush J.A. Phalangeal morphology of the Paromomyidae (?Primates, Plesiadapiformes): the evidence for gliding behavior reconsidered // AJPhA, 1999, V.109, pp.397–413.
Hand S.J., Novacek M., Godthelp H. et Archer M. First Eocene bat from Australia // Journal of Vertebrate Paleontology, 1994, V.14, pp.375–381.
Hartenberger J.-L. Hypothèse paléontologique sur l’origine des Macroscelidea (Mammalia). Comptes Rendus de l’Académie des Sciences, Serie II, 1986, V.302, pp.247–249.
Henry A.G., Brooks A.S. et Piperno D.R. Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium) // PNAS, 2011, V.108, № 2, pp.486–491.
Hooker J.J. A primitive emballonurid bat (Chiroptera, Mammalia) from the earliest Eocene of England // Palaeovertebrata, 1996, V.25, pp.287–300.
Hooker J.J., Russell D.E. et Phélizon A. A new family of Plesiadapiformes (Mammalia) from the Old World Lower Paleogene // Palaeontology, 1999, V.42, № 3, pp.377–407.
Hu Y., Meng J., Wang Y., Li Ch. Large Mesozoic mammals fed on young dinosaurs // Nature, 2005, V.433, № 7022, pp.149–152.
Hutcheon J.M., Kirsch J.A. et Pettigrew J.D. Base-compositional biases and the bat problem. III. The questions of microchiropteran monophyly // Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences, 1998, V.353, № 1368, pp.607–617.
Ji Q., Luo Zh.-X., Yuan Ch.-X., Wible J.R., Zhang J.-P. et Georgi J.A. The earliest known eutherian mammal // Nature, 2002, V.416, pp.816–822.
Ji Q., Luo Zh.-X., Yuan Ch.-X. et Tabrum A.R. A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals // Science, 2006, V.311, № 5764, pp.1123–1127.
Koenigswald von W. et Schierning H.-P. The ecological niche of an extinct group of mammals, the early Tertiary apatemyids // Nature, 1987, V.326, pp.595–597.
Kay R.F. et Cartmill M. Cranial morphology and adaptations of Palaeehthon naeimienti and other Paromomyidae (Plesladapoldea,? Primates), with a description of a new genus and species // JHE, 1977, V.6, pp.19–53.
Krause D.W. Were paromomyids gliders? Maybe, maybe not // JHE, 1991, V.21, pp.177–188.
Ladevèze S., Muizon de Ch., Beck R.M.D., Germain D. et Cespedes-Paz R. Earliest evidence of mammalian social behaviour in the basal Tertiary of Bolivia // Nature, 2011, V.474, pp.83–86.
Lilly J.C. The cetacean brain // Oceans, 1977, V.10, pp.4–7.
Lofgren D.L. The Bug Creek problem and the Cretaceous-Tertiary transition at McGuire Creek, Montana // University of California publications in geological sciences, 1995, V.140, pp.1-185.
Luo Zh.-X., Crompton A.W., Sun A.-L. A new Mammaliaform from the Early Jurassic and evolution of Mammalian characteristics // Science, 2001, New Series, Vol. 292, № 5521, pp.1535–1540.
Luo Zh.-X. et Wible J.R. A Late Jurassic Digging Mammal and Early Mammal Diversification // Science, 2005, V.308, pp.103–107.
Luo Z.-X.,Yuan Ch.-X., Meng Q.-J. et Ji Q. A Jurassic eutherian mammal and divergence of marsupials and placentals // Nature, 2011, V.476, pp.442–445.
MacPhee R.D.E., Cartmill M. et Rose K. D. Craniodental Morphology and Relationships of the Supposed Eocene Dermopteran Plagiomene (Mammalia) // Journal of Vertebrate Paleontology, 1989, V.9, № 3, pp.329–349.
Marshall L.G. et Muizon de C. The Dawn of the Age of Mammals in South America // National Geographic Research, 1988, V.4, № 1, pp.23–55.
Martin J.E., Case J.A., Jagt J.W.M., Schulp A.S. et Mulder E.W.A. A New European Marsupial Indicates a Late Cretaceous High-latitude Transatlantic Dispersal Route // Journal of Mammalian Evolution, 2005, V.12, № 3/4, pp.495–511.
Martin Th. Postcranial anatomy of Haldanodon exspectatus (Mammalia, Docodonta) from the Late Jurassic (Kimmeridgian) of Portugal and its bearing for mammalian evolution // Zoological Journal of the Linnean Society, 2005, V.145, pp.219–248.
Meng J., Hu Y., Wang Y., Wang X. et LiCh.A Mesozoic gliding mammal from northeastern China // Nature, 2006, V.444, pp.889–893.
Meredith R.W., Janecka J.E., Gatesy J., Ryder O.A., Fisher C.A., Teeling E.C., Goodbla A., Eizirik E., Simão T.L.L., Stadler T., Rabosky D.L., Honeycutt R.L., Flynn J.J., Ingram C.M., Steiner C., Williams T.L., Robinson T.J., Burk-Herrick A., Westerman M., Ayoub N.A., Springer M.S., Murphy W.J. Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification // Science, 2011, V.334, pp.521–524.
Mindell D.P., Dick Ch.W. et Baker R.J. Phylogenetic relationships among megabats, microbats,and primates(molecular systematics // PNAS, 1991, V.88, pp.10322-10326.
Moffat A.S. New fossils and a glimpse of evolution // Science, 2002, V.295, pp.613–615.
Muizon de C. et McDonald H.G. An aquatic sloth from the Pliocene of Peru // Nature, 1995, V.375, № 6528, pp.224–227.
Orkin J.D. et Pontzer H. The narrow niche hypothesis: gray squirrels shed new light on primate origins // AJPhA, 2011, V.144, pp.617–624.
Penkrot T.A., Zack Sh.P., Rose K.D. et Bloch J.I. Postcranial Morphology of Apheliscus and Haplomylus (Condylarthra, Apheliscidae): Evidence for a Paleocene Holarctic Origin of Macroscelidea // Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay. Eds.: E.J. Sargis et M. Dagosto. 2008, pp.73-106.
Pettigrew J.D. Flying primates? Megabats have the advanced pathway from eye to midbrain // Science, 1986, V.231, № 4743, pp.1304–1346.
Pettigrew J.D., Maseko B.C. et Manger P.R. Primate-like retinotectal decussation in an echolocating megabat, Rousettus aegyptiacus // Neuroscience, 2008, V.153, № 1, pp.226–231.
Pettigrew J.D., Jamieson B.G., Robson S.K., Hall L.S., McAnally K.I. et Cooper H.M. Phylogenetic relations between microbats, megabats and primates (Mammalia: Chiroptera and Primates) // Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences, 1989, V.325, № 1229, pp.489–559.
Prasad G.V.R., Verma O., Sahni A., Parmar V. et Khosla A. A cretaceous hoofed mammal from India // Science, 2007, V.318, № 5852, p.937.
Rinderknecht A. et Blanco R.E. The largest fossil rodent // Proceedings of the Royal Society, 2008, Ser.B, V.275, № 1637, pp.923–928.
Rydell J. et Speakman J.R. Evolution of nocturnality in bats — potential competitors and predators intheir early history. Biological Journal of the Linnaean Society, 1995, V.54, pp.183–191.
Sánchez-Villagra M.R., Aguilera O. et Horovitz I. The Anatomy of the World's Largest Extinct Rodent // Science, 2003, V.301, № 5640, pp.1708–1710.
Seiffert E.R. The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt // Acta Palaeontologica Polonica, 2010, V.55, pp.599–616.
Simmons N.B., Seymour K.L., Habersetzer J. et Gunnell G.F. Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation // Nature, 2008, V.451, pp.818–822.
Simons E.L., Holroyd P.A. et Bown Th.M. Early tertiary elephant-shrews from Egypt and the origin of the Macroscelidea // PNAS, 1991, V.88, pp.9734–9737.
Simpson G.G. The Tertiary lorisiform primates of Africa // Bulletin of the Museum of Comparative Zoology, 1967, V.136, pp.39–62.
Simpson G.G. The Tiffany fauna, Upper Paleocene.I. — Multituberculata, Marsupialia, Insectivora and?Chiroptera // American Museum Novitates, 1935, № 795, pp.1-19.
Smith Th., De Bast E. et Sigé B. Euarchontan affinity of Paleocene Afro-European adapisoriculid mammals and their originin the late Cretaceous Deccan Traps of India // Naturwissenschaften, 2010, V.97, pp.417–422.
Speakman J.R. The evolution of flight and echolocation in bats: another leap in the dark // Mammal Rev., 2001, Volume 31, No. 2, 111–130.
Sussman R.W. Primate Origins and the Evolution of Angiosperms // American Journal of Primatology, 1991, V.23, pp.209–223.
Szalay F.S. Mixodectidae, Microsyopidae, and the insectivore-primate transition // Bulletin of the American Museum of Natural History, New York, 1969, V.140, article 4, pp.193–330.
Szalay F.S. et Delson E. Evolutionary history of the Primates. New York, Academic Press, 1979.
Tabuce R., Coiffait B., Coiffait Ph.-E., Mahboubi M. et Jaeger J.-J. A new genus of Macroscelidea (Mammalia) from the Eocene of Algeria: a possible origin for elephant-shrews // Journal of Vertebrate Paleontology, 2001, V.21, № 3, pp.535–546.
Van Valen L. New Paleocene insectivores and insectivore classification // Bulletin of the American Museum of Natral History, New York, 1967, V.135, article 5, pp.217–284.
Veselka N., McErlain D.D., Holdsworth D.W.,EgerJ.L., Chhem R.K., Mason M.J., Brain K.L., Faure P.A. et Brock Fenton M. A bony connection signals laryngeal echolocation in bats // Nature, 2010, V.463, pp.939–942.
Walker A.C. True affinities of Propotto leakeyi Simpson, 1967 // Nature, 1969, V.223, pp.647–648.
Wells N.A. et Gingerich P.D. Review of Eocene Anthracobunidae (Mammalia, Proboscidea) with a new genus and species, Jozaria palustris, from the Kuldana Formation of Kohat (Pakistan) // Contributions from the Museum of Paleontology, The University of Michigan, 1983, V.26, № 7, pp.117–139.
West R.M. et Dawson M.R. Vertebrale Paleontology and the Cenozoic History of the North Atlantic Region // Polarforschung, 1978, V.48, № 1/2, pp.103–119.
Wible J.R., Rougier G.W., Novacek M.J. et Asher R.J. Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary // Nature, 2007, V.447, pp.1003–1006.
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги