Часть I Первые наброски
Глава 2
От одной клетки к множеству
Я широк, я вмещаю в себе множество разных людей.
Я широк, я вмещаю в себе множество разных людей.
Величайшая ирония биологии в том, что человеческий организм – одна из самых сложных сущностей во Вселенной,[3] – имеет очень простое начало. Взрослый человек состоит из миллионов миллионов клеток. Их в десять раз больше, чем звезд в нашей галактике, или, если привести более земное сравнение, в десять раз больше, чем песчинок на небольшом пляже. Все это огромное количество клеток не свалено как попало, а организовано в сложные структуры, и, несмотря на многовековую историю анатомических исследований, мы все еще не знаем всех деталей их строения. Существуют сотни типов клеток, и у каждого своя функция и свой образ жизни, при этом клетки каждого конкретного типа появляются и обновляются в нужном количестве и в нужном месте. Вся эта сложнейшая конструкция развивается из одной оплодотворенной яйцеклетки. И скромной клетки, внешне лишенной каких-либо особых признаков. И уже с самого начала постепенное усложнение человеческого организма протекает в режиме самоорганизации: ему приходится действовать самостоятельно, без какой-либо посторонней помощи.
Первый шаг к усложнению – переход от одной клетки к множеству. Это обусловлено необходимостью одновременного протекания большого количества различных процессов. Например, сейчас вы дышите, перевариваете пищу и выводите из организма вредные вещества; ваши волосы растут, кожа обновляется, кровь фильтруется, иммунная система борется с болезнетворными бактериями, температура тела регулируется; вы слушаете, читаете и думаете, а теперь, когда вы дочитали это предложение до конца, возможно, также прислушиваетесь к своему организму. Эти процессы, как и многие другие, приводятся в действие с помощью особых белков и биохимических механизмов. Многие из этих процессов никак не могут протекать в одном и том же месте. Например, представим, что в одном и том же месте организма матери вырабатывается молоко для ребенка и тут же переваривается то молоко, которое она только что выпила с чаем. Есть и другие примеры процессов, несовместимых по более сложным причинам: из-за особенностей строения белков или функционирования генов.
Сложные организмы справляются с этой проблемой благодаря компартментализации – принципу разделения процессов в пространстве. Тело состоит из органов, имеющих определенные функции, органы – из специализированных тканей, а ткани – из разных типов клеток. Однако внутри клетки большая часть молекул постоянно находится в движении, из-за чего трудно достичь одновременного выполнения многих операций. Компартментализация существует и на уровне клетки. В главе 8, посвященной перемещению клеток эмбриона и изменению их взаимного расположения, мы обсудим, как разные части клетки могут выполнять несколько разные функции. Однако «многозадачность» клетки имеет свои пределы. Поэтому мы будем считать клетку базовой единицей, которая одновременно выполняет лишь одно-два дела. Именно поэтому многообразие типов клеток является необходимым шагом к созданию сложного организма.
Механизмы, за счет которых одна клетка превращается в две, а потом и во множество клеток, не только принципиально важны для эмбрионального развития, но и ярко демонстрируют возможности самоорганизации. Простые маленькие молекулы могут самоорганизовываться в крайне сложные структуры, имеющие гораздо больший пространственный масштаб, чем сами молекулы, причем без какого бы то ни было предварительного плана. Это краеугольный камень, лежащий в основе понимания развития эмбриона. Поэтому в этой главе мы подробно остановимся на механизмах деления клетки, а в дальнейшем будем принимать их как данность.
Оплодотворенная яйцеклетка, с которой начинается развитие человека, необычно велика. Она достигает десятой доли миллиметра в диаметре и видна даже невооруженным глазом. Большинство клеток организма намного меньше: примерно сотая доля миллиметра в диаметре и тысячная доля объема яйцеклетки. Это означает, что оплодотворенная яйцеклетка может превратиться в многоклеточный эмбрион просто поделившись на две, затем на четыре, на восемь и так далее, без перерывов на рост. Такой тип деления клетки – дробление – очень удобен для эмбриона, так как позволяет отложить проблему питания, обеспечивающего энергию для роста, на потом, а именно на этап, когда эмбрион уже станет многоклеточным и сможет выделить для переработки пищи специализированные части тела.
Если расти не нужно, процесс деления сводится к распределению молекул (например, белков) поровну между дочерними клетками. Суть деления при неизменном объеме заключается в сохранении концентрации белков и питательных веществ. Ярким исключением из этого правила является молекула ДНК: в исходной клетке сорок шесть хромосом (двадцать три от матери и двадцать три от отца), и каждая новая клетка должна содержать такое же их число. Поэтому хромосомы должны копироваться (реплицироваться) перед каждым делением клетки. Более того, должны существовать специальные системы, гарантирующие «честное» распределение реплицированных хромосом по дочерним клеткам – каждая из них должна получить не сорок шесть любых хромосом, а по одной копии каждой хромосомы от отца и по одной – от матери. Система, которая обеспечивает эту нелегкую задачу, является одной из основных систем клеток животных и растений и существует уже около 2,5 млрд лет. И всего пару миллионов лет назад появились существа, которые в принципе способны понять, как она работает.
Копирование ДНК – самая простая и самая древняя часть процесса, ей уже как минимум 3,5 млрд лет. Она основана на том, что молекулы ДНК существуют в виде пары нуклеотидных цепей (иногда их называют «нитями»). Аденину на одной цепи всегда соответствует тимин на другой цепи, а цитозину – гуанин. Это строгое правило, связанное с химической структурой нуклеотидов, означает, что каждая цепочка содержит всю необходимую информацию о последовательности соседней цепи. Репликация ДНК начинается с того, что ферментный комплекс отделяет две материнских цепи друг от друга. Затем он собирает новую цепь для каждой из них, соединяя нуклеотиды в порядке исходной цепи. Каждая новая цепь соединяется со старой, которая служила для нее матрицей. В результате получаются две молекулы ДНК вместо одной. То есть происходит репликация ДНК. Белки, в которые завернута ДНК, добавляются сразу же после копирования.
После репликации сорок шесть хромосом одноклеточного зародыша должны быть распределены так, чтобы каждой дочерней клетке непременно досталось по одной копии каждой материнской хромосомы и каждой отцовской хромосомы. Этот процесс можно разбить на несколько этапов: 1) определение центров двух дочерних клеток; 2) выравнивание всех скопированных хромосом между этими центрами; 3) «растаскивание» копий – по одному экземпляру каждой пары отходит каждой дочерней клетке; 4) отделение дочерних клеток друг от друга. Каждый из этих этапов включает в себя координированные действия в пространственном масштабе, значительно превышающем размеры вовлеченных в них молекул. При этом весь процесс должен протекать без отклонений, несмотря на то что точное расположение основных компонентов (хромосом, например) будет постоянно меняться. Поэтому все эти этапы в значительной степени полагаются на адаптивную самоорганизацию и могут служить отличным примером для иллюстрации этого принципа.
Первая проблема заключается в выявлении центров новых дочерних клеток. Проще всего понять, как определяется центр в типичной взрослой клетке, которая не собирается делиться, а просто находится в состоянии покоя. На первый взгляд проблема кажется простой. Однако при более детальном рассмотрении все усложняется. Клетки вообще не имеют четкой формы: она зависит от их окружения. Это исключает любой заранее заготовленный план. Диаметр типичной клетки человека – примерно одна сотая миллиметра – кажется нам небольшим, ведь мы состоим из миллионов клеток. Однако это в тысячу раз больше, чем длина обычной молекулы белка. Тем не менее комплексы белков каким-то образом находят центр клетки. Это все равно что запустить в Альберт-Холл[4] глухих людей с завязанными глазами и попросить их найти его середину.
Клетка нашла крайне остроумный способ решения этой проблемы. Он хорошо иллюстрирует, насколько важны могут быть тривиальные детали биохимических процессов для функционирования клетки. «Звездой» всего действа является белок тубулин, молекулы которого связываются друг с другом и образуют длинные структуры – микротрубочки. Одна из особенностей сборки молекул тубулина заключается в том, что объединение нескольких молекул тубулина для образования новой микротрубочки – событие маловероятное, а процесс присоединения молекулы тубулина к уже существующей микротрубочке, то есть ее удлинение, протекает относительно легко. Поэтому микротрубочки, как правило, не образуются спонтанно, но после того, как они образовались, они способны к спонтанному росту.
Вторая особенность биохимии тубулина заключается в том, что каждая отдельно взятая молекула может находиться в одном из двух состояний, «свежем» или «несвежем».[5] «Свежие» молекулы медленно превращаются в «несвежие». Только «свежие» молекулы могут присоединяться к концам существующих микротрубочек. Концы микротрубочек устойчивы только тогда, когда они созданы из «свежего» тубулина (если концы остаются «свежими», то не имеет значения, утратит ли тубулин «свежесть» по всей длине микротрубочки).[6] Если на конце микротрубочки «несвежий» тубулин, то конец начинает распадаться, и распад продолжается до тех пор, пока микротрубочка не присоединит стабильный «свежий» тубулин. Учитывая, что тубулин вдали от концов микротрубочки, скорее всего, был в ее составе дольше, чем тот, из которого состоит ее конец, эти «внутренние» молекулы, скорее всего, давно испортились, и «свежего» тубулина, способного предотвратить разрушение микротрубочки, не осталось. В таком случае микротрубочка распадется. Единственный способ избежать распада, не прибегая к помощи других молекул, – это быстрый рост, при котором «свежий» тубулин присоединяется к концу микротрубочки быстрее, чем разрушается «несвежий». Таким образом, микротрубочки либо быстро растут, либо катастрофически быстро распадаются. Существует постоянная вероятность разрушения, а это означает, что длинных трубочек всегда меньше, чем коротких. Эта особенность имеет непосредственное отношение к механизму нахождения клеточных центров.
Молекулы тубулина редко самопроизвольно объединяются в новые микротрубочки, и поэтому в клетке есть особые комплексы белков, которые могут катализировать этот процесс. Эти комплексы располагаются в ключевом месте клетки, а именно в центросоме, от которой микротрубочки расходятся радиально, как спицы от ступицы колеса.[7] Пока они растут достаточно быстро для того, чтобы тубулин на их концах оставался свежим, микротрубочки будут удлиняться по направлению к периферии клетки. Существует две теории о том, как они помогают центросоме попасть в центр клетки. Они основаны на экспериментальных данных, полученных при исследовании различных организмов. Еще не ясно, какая из них справедлива для эмбрионов человека; не исключено, что обе. Одна из теорий связана с отталкиванием, а другая – с подтягиванием.
Механизм отталкивания[8] основан на способности растущих микротрубочек отталкиваться от внутренней поверхности клеточной мембраны. Если центросома располагается близко к одной из сторон клетки, даже короткие микротрубочки смогут достичь поверхности мембраны и оттолкнуться от нее. В результате центросома отдаляется от этой стороны. Противоположной стороны клетки достигают только очень длинные микротрубочки, но они по упомянутым выше причинам встречаются редко. А поскольку таких микротрубочек меньше, они будут слабее отталкивать центросому от этой стороны клетки. Такое неравномерное распределение сил оттолкнет центросому от ближайшей мембраны, и она займет стабильное положение только тогда, когда силы отталкивания придут в равновесие. Равновесие же наступает тогда, когда центросома находится на одинаковом расстоянии от всех сторон: другими словами, в центре клетки (рис. 4, а). Ученые поместили центросомы в искусственно изготовленную «ячейку» и доказали, что им удалось найти центр «ячейки» с помощью механизма отталкивания.[9]
Механизм подтягивания[10],[11],[12] основан на действии небольших моторных белков, распределенных по всей клетке. Они могут связываться с микротрубочками и перемещаться по ним в сторону центросомы. Продвигаясь к центросоме, каждый из этих белков генерирует силу, которая слегка тянет микротрубочку в противоположном направлении, смещая ее в сторону клеточной мембраны. Так человек, идущий вперед по лодке, толкает ее, смещая назад. Чем длиннее микротрубочка, тем больше моторных белков могут связаться с ней и тем сильнее они за нее тянут в нужном направлении.[13] Таким образом, если центросома ближе к одной стенке клетки, чем к другой, моторные белки сильнее всего тянут за длинные микротрубочки, направленные к дальней стороне, смещая центросому к центру клетки (рис. 4, б). Тщательные исследования оплодотворенных яйцеклеток просто устроенных организмов (таких, как морской еж или нематода) показали, что важную роль в этих клетках играет именно механизм подтягивания. Например, если часть микротрубочек разрезать с помощью лазера, центросома отскочит назад так, словно бы ее удерживали на месте натянутые микротрубочких.[14] Не исключено, что в некоторых клетках работают оба механизма: сильное натяжение длинных микротрубочек «кнаружи» еще больше снижает их способности толкать центросому и усиливает дисбаланс сил отталкивания.

Рис. 4. Две теории, объясняющие, как центросома находит центр клетки с помощью микротрубочек. В модели отталкивания (а) микротрубочки отталкиваются от клеточной мембраны. Коротких трубочек всегда больше, чем длинных. Значит, центросома отталкивается сильнее от ближайшей клеточной мембраны. В модели подтягивания (б) моторные белки, распределенные по всей клетке, прикрепляются к длинным микротрубочкам. Коротких микротрубочек много со всех сторон центросомы, а длинные могут сформироваться только с удаленной от мембраны стороны. Поэтому центросому тянут к дальней стороне клетки и она удаляется от ближайшей мембраны
Однако какой бы из этих механизмов ни использовался клетками эмбриона человека, результат будет одним и тем же: центросома отцентрирует себя автоматически, хотя ни один из задействованных в этом процессе компонентов клетки не «знает» ее формы и не руководствуется для поиска центра какой-либо системой координат. Система организует себя сама. За такую самоорганизующуюся систему, способную адаптироваться практически к любым условиям, приходится расплачиваться энергией, необходимой для сборки микротрубочек и работы моторных белков; высокие энергетические затраты вообще характерны для адаптивной самоорганизации.
При клеточном делении необходимо определить не просто центр одной клетки, а те точки, которые будут центрами двух дочерних клеток, чтобы хромосомы переместились в правильном направлении. К счастью, клетка может при помощи того же механизма определить и два центра; все, что для этого нужно, – это две центросомы.
Центросомы состоят из «облака» белков вокруг пары соединенных вместе трубчатых структур, содержащих тубулин.[15] Эта пара является организационным центром для материала центросомы. В процессе подготовки к делению клетки трубочки отсоединяются друг от друга, и каждая из них сразу же становится матрицей для сборки недостающего партнера. Таким образом, через некоторое время по соседству будут располагаться уже две пары трубчатых структур. Каждая из них организует вокруг себя центросомный материал и инициирует образование новых микротрубочек, радиально расходящихся от центросомы. В клетке с двумя центросомами радиальные микротрубочки одной системы «наталкиваются» на микротрубочки другой. В модели отталкивания микротрубочки одной системы будут отталкиваться от микротрубочек другой системы, точно так же как от клеточной мембраны. Присутствие второй центросомы и второй системы микротрубочек создает «ложное впечатление» о том, насколько близко каждая из центросом находится к клеточной мембране. Поэтому каждая из центросом оказывается не в центре клетки, а на максимальном удалении от другой центросомы (рис. 5). Аналогичным образом, в модели подтягивания с каждая система, состоящая из центросомы и микротрубочек, служит щитом для другой и не дает утащить центросому к дальней стороне клетки. Оба механизма, которые в клетках человека могут работать одновременно, будут иметь один и тот же эффект: ни одна центросома не будет находиться в центре клетки. Вместо этого они займут положение примерно посередине между истинным центром и периферией клетки (рис. 5). Таким образом, две центросомы определяют будущие центры двух новых клеток, которые образуются при делении материнской клетки. Опять же, это происходит «автоматически» – участники процесса ничего не «знают» о форме клетки.

Рис. 5. Когда в клетке две центросомы, взаимодействие микротрубочек приводит к их расхождению
Радиальные микротрубочки, отходящие от центросом, служат не только для определения будущих клеточных центров. Они также способствуют расхождению двойного набора хромосом, чтобы каждая новая клетка получила их полный комплект. Чтобы это произошло, микротрубочкам сначала нужно соединиться с хромосомами. Опять же, ни одному из компонентов не требуется «знание» о положении других участников процесса. Система снова пользуется нестабильностью микротрубочек, которые проходят периоды сборки и катастрофического распада. Незащищенные концы микротрубочек особенно подвержены разборке, но, если они будут окружены особыми белками, способными связываться с микротрубочками, ситуация немного стабилизируется. В каждой хромосоме есть один специальный участок, содержащий такие белки, поэтому любая растущая микротрубочка может случайно «наткнуться» на этот участок и получить защиту.[16] Таким образом, система, в которой микротрубочки расходятся случайным образом и не погибают только в том случае, если сталкиваются с хромосомой, приводит к тому, что все хромосомы оказываются «заякорены» при помощи микротрубочек.
Простейшей связи микротрубочек с хромосомами достаточно, чтобы каждая хромосома переместилась к центрам дочерних клеток, но для деления клетки требуется нечто большее. Одна копия хромосомы, скажем 9-й хромосомы отца, должна присоединиться к микротрубочке одной из центросом, а вторая ее копия должна присоединиться к микротрубочкам другой центросомы. Таким образом, каждая из них направится в свою клетку. Это достигается еще одним воздействием на устойчивость микротрубочек. Две копии каждой хромосомы, образовавшиеся в процессе репликации ДНК, соединены с помощью специальных белковых комплексов. Эти комплексы находятся в состоянии механического напряжения, когда два набора микротрубочек и связанные с ними моторные белки играют в «перетягивание каната» и пытаются растащить две копии хромосомы к противоположным центросомам. В этой ситуации генерируется сигнал, который приводит к более надежной стабилизации микротрубочек, чем в отсутствие механического напряжения.[17] Если обе копии хромосомы прикреплены к микротрубочкам из одной системы, напряжения не возникнет, и микротрубочки долго не продержатся. Если они прикреплены к микротрубочкам разных центросом, которые пытаются оттащить их друг от друга, сигнал стабилизации достаточно сильный, и у микротрубочек гораздо больше шансов на выживание. Поэтому система постоянно меняется, продолжает находиться в режиме «поискового поведения», пока не находит то самое состояние, в котором копии хромосом будут растаскиваться в противоположных направлениях.[18] Этот способ требует больших энергетических затрат, но зато действует автоматически и позволяет справиться даже с дополнительными хромосомами, появившимися в клетке в процессе эволюционных изменений или в ходе экспериментов.
Когда все хромосомы выстроились в линию, клетка может перейти к следующему этапу деления, на котором белки, связывавшие сестринские копии хромосомы, освобождают их, позволяя смещаться к противоположным сторонам клетки. Процесс не начнется до тех пор, пока все хромосомы не выстроены правильно, иначе дочерние клетки получат неправильное число хромосом и лишатся важных генов. Поэтому должна существовать система, предотвращающая преждевременное расхождение. В очередной раз используется способность белков, соединяющих копии хромосом, «ощущать» напряжение, возникающее, когда копии хромосомы крепятся к микротрубочкам из двух разных систем. Когда напряжение отсутствует, белковые комплексы продолжают генерировать сигнал, небольшую молекулу, которая может распространиться по клетке и блокировать переход к следующей стадии клеточного деления. Она как будто кричит «Не сейчас!» на языке биохимии. До тех пор пока все пары хромосомы не будут прикреплены к правильным микротрубочкам и не будут подвергаться растаскиванию, сигналы будут продолжаться и клетка будет ждать. Только тогда, когда натяжение всех белковых комплексов будет отрегулировано, сигналы смолкнут и можно будет начинать следующий этап деления. Опять же, эта система может приспособиться под любое количество хромосом.
Когда все хромосомы выстроились в середине веретена деления, а сигналы «Не сейчас!» затихли, клетка готова к следующему этапу. Белковые комплексы, которые соединяют пары копий хромосом, отпускают их, позволяя моторным белкам свободно подтягивать хромосомы по микротрубочкам к соответствующим центросомам.[19] После того как все хромосомы распределились, еще одна автоматическая система размещает кольцо сократительных белков по «экватору» клетки между полюсами, положение которых определяют центросомы. Эти белки могут скользить относительно друг друга, образуя затягивающийся «пояс» клетки. Этот «пояс» будет затягиваться до тех пор, пока исходная клетка не разделится окончательно на две новые.
В целом описанные выше системы могут показаться очень сложными и запутанными. Однако, если в их устройстве разбираться последовательно, компонент за компонентом, они оказываются очень простыми. У каждого белка своя простая задача. Способность системы выполнять сложные задачи, такие как обнаружение и разделение сестринских хромосом, независимо от их исходного положения в клетке, обеспечивается взаимодействием простых компонентов системы, а не их усложнением. В частности, она связана с тем, что существует обратная связь между поведением компонентов системы и получаемой ими информации о состоянии системы как целого, о том, на какой стадии того или иного процесса она находится в данный момент (например, о том, все ли хромосомы заняли нужное положение). Использование простых компонентов, увязанных в единое целое за счет системы тесных обратных связей, вообще характерно для живой материи, и в этой главе я углубился в детали именно затем, чтобы показать, как системы «глупых» биологических молекул могут организоваться для решения серьезных задач, иначе говоря, чтобы показать, как из простого возникает сложное.
Системы, которые запускали первое деление клетки, продолжают работать в клетках эмбриона, и теперь мы больше не будем обсуждать их в таких деталях. Это вообще типично для биологии: если механизм работает, его можно использовать многократно, иногда с незначительными поправками, в процессе всего эмбрионального развития. Почти сразу после того, как первое деление клетки завершено, каждая из двух образовавшихся клеток начинает копировать хромосомы и делиться. Получается эмбрион из четырех клеток. Синхронные клеточные деления продолжаются некоторое время, но рано или поздно синхронность деления разных клеток теряется, и со стадии примерно шестнадцати клеток число клеток эмбриона начинает отклоняться от значения «два в степени n». Как правило, клетки раннего эмбриона, получившиеся в ходе дробления, расположены довольно свободно относительно друг друга. Иногда, примерно один раз на тысячу двести случаев, клетки распадаются на два отдельных комка. Из каждого комка получится отдельный эмбрион, у которого будет собственная плацента и плодные оболочки. Это один из трех вариантов рождения однояйцевых близнецов, и на него приходится около трети всех случаев. Тот факт, что эмбрион может разделиться и дать начало двум младенцам, говорит нам об очень важной особенности развития: все клетки должны быть способны создать любую часть тела, и нет одной специальной клетки, которая отвечает, например, за создание головы или детерминирована как клетка головы. Если бы клетки с самого начала были разными, если бы одна или несколько клеток были детерминированы к формированию конкретной части тела или если бы одна клетка отвечала за весь процесс, то разделение эмбриона привело бы к тому, что по меньшей мере у одной из половинок не оказалось бы жизненно важных клеток и ее развитие было бы обречено на провал. Тем не менее тысячи живущих в каждой стране однояйцевых близнецов являются красноречивым свидетельством равных возможностей клеток на ранних этапах дробления.
Итак, эмбрион достигает стадии примерно шестнадцати клеток. Это значит, что у него уже достаточно клеток, чтобы изменять форму тела и чтобы начать клеточную дифференцировку, то есть сделать клетки разными. Прелюдия окончена, и начинается основная работа по эмбриональному развитию.
Глава 3
Как создаются различия
Искреннее расхождение во мнениях – хороший признак прогресса.
Искреннее расхождение во мнениях – хороший признак прогресса.
Суть раннего дробления можно выразить одной фразой: «умножение без изменения». Количество клеток растет в геометрической прогрессии, но каждая новая клетка идентична всем остальным. Действительно, во время первых нескольких делений клетки даже не пользуются собственными генами, а полагаются на запас молекул, помещенных материнским организмом в яйцеклетку и поровну распределяемых при делении между дочерними клетками.[20] Ранние этапы развития полностью посвящены делению. Это имеет смысл: чем больше клеток, тем легче будет в нужный момент создать с их помощью организм. Однако дробление может продолжаться только до определенного момента, так как образующиеся клетки имеют вдвое меньший объем, чем их предшественники. Через некоторое время размер клеток приблизится к минимальному, и им понадобятся паузы для роста между новыми этапами деления. Для роста же необходимы питательные вещества, а чтобы их получить, нужно приложить дополнительные усилия. Это, в свою очередь, означает, что какие-то из клеток должны специализироваться на доставке пищи другим. В эмбрионах многих птиц и рептилий питание осуществляется за счет запасенного в яйце желтка. В случае млекопитающих питательные вещества поступают непосредственно от матери, но общее правило остается неизменным: рано или поздно дробление прекращается и начинается специализация клеток.
Специализация подразумевает, что клетки теряют идентичность. Теперь у разных типов клеток разные функции. На этом этапе эмбрион сталкивается с очень важной проблемой. Создание различий означает необходимость создания нового порядка и новой информации. Увеличение количества информации хорошо иллюстрируется таким примером: для описания асимметричного объекта нам требуется больше слов или, скажем, математических символов, чем для описания симметричного объекта. Так, описать форму кружки с ручкой сложнее, чем описать форму, скажем, стакана. У многих «низших» существ, например насекомых, необходимую информацию обеспечивает мать. Она закладывает пространственную информацию в яйцеклетку в виде градиентов концентрации определенных молекул. Таким образом, во время дробления разные клетки эмбриона наследуют разное количество этих молекул. Затем клетки используют эти различия для выбора пути развития. Этот негенетический метод передачи чрезвычайно важной информации из поколения в поколение очень эффективен, но, насколько мы знаем, у человеческих эмбрионов он не работает, и одна часть яйцеклетки идентична другой.[21] Создание отличительных особенностей – то есть новой информации – представляет собой серьезную логическую задачу для человеческого эмбриона. Как создать закономерность, которой раньше не было? У этой проблемы очень изящное решение: эмбрион получает информацию исходя из законов геометрии.
Пока эмбрион состоял из нескольких клеток, каждая из них занимала достаточно большую долю в общем объеме. Часть поверхности каждой клетки была обращена в окружающую среду. Когда в ходе дробления получается тридцать две или шестьдесят четыре клетки, уже достаточно мелкие по сравнению с размерами сферического зародыша, некоторые из них оказываются окружены со всех сторон другими клетками. Другие все еще имеют контакт с внешней средой – примерно одна шестая их мембраны обращена наружу. Клетки «чувствуют», окружены ли они другими клетками или какая-то их часть соприкасается только с жидкостью. Эту информацию они используют для того, чтобы определиться с дальнейшими действиями. Клетки, имеющие «свободную поверхность», активируют ряд ранее неактивных генов и образуют первую ткань эмбриона – трофэктодерму. Клетки, окруженные другими клетками, эти гены не активируют. Использование в качестве источника информации простого физического признака – наличия свободной поверхности – избавляет эмбрион от необходимости иметь заранее подготовленный пространственный план. Кроме того, клеткам не нужно «знать» свое точное расположение в эмбрионе. Им нужно только определить, есть у них свободные участки поверхности или нет.
Задача трофоэктодермы заключается только в создании структур для структурной поддержки и питания эмбриона. Частью ребенка она никогда не станет.[22] В первую очередь трофэктодерма перекачивает в эмбрион жидкость. Эта жидкость накапливается, что приводит к образованию обширной полости (рис. 6). Формирование этой полости, в свою очередь, приводит к тому, что внутренние клетки группируются на внутренней поверхности трофэктодермы в виде эксцентрично расположенного комка, который называется «внутренняя клеточная масса». Если судить только по внешнему виду, эти клетки кажутся менее интересными, чем активные клетки трофэктодермы, постоянно занятые питанием, накачкой жидкости, внедрением в материнский организм. Однако именно из внутренней клеточной массы и формируется ребенок. Иногда эта масса делится на два комочка, каждый из которых развивается, формируя в итоге человеческий организм. Такие близнецы, как и те, о которых рассказывалось в главе 2, будут генетически идентичны, но на этот раз они будут расти в одной общей трофэктодерме. В то же время у каждого из близнецов будет свой желточный мешок и амниотическая полость (см. далее). Таким образом, они надежно изолированы друг от друга. Это самый распространенный механизм появления однояйцевых близнецов, на его долю приходится две трети всех случаев. (Третий, крайне редкий вариант появления близнецов будет описан несколько позже.)
К тому времени, когда клетки трофэктодермы закачают внутрь эмбриона объем жидкости, достаточный для создания полости, и создают вздувшуюся структуру, эмбрион должен покинуть фаллопиеву трубу матери, где произошло оплодотворение, и переместиться в матку. Этот орган в норме как бы немного «сдут»: его стенки слипаются, как снятая с руки резиновая перчатка. Особенно «сдувшейся» матка выглядит через неделю после овуляции. Поэтому весьма вероятно, что ранний эмбрион, попав в матку, вскоре столкнется с ее внутренней поверхностью. Когда это произойдет, он закрепляется на ней, используя специальный набор адгезивных молекул. После прикрепления клетки эмбриона образуют новые белки, которые позволяют им внедриться между клетками стенки матки.[23] В течение нескольких часов «пальчики» клеток эмбриона проникнут в ткани матери, и образуется плацента. Многие клетки матки при этом разрушаются, а их остатки становятся пищей для эмбриона. Реакция матери на эту атаку приводит к дальнейшему отмиранию ткани. Таким образом, через десять дней после оплодотворения формируется полость, в которой легко умещается эмбрион. У людей и некоторых животных внутренняя выстилка матки обрастает эмбрион, скрывая место имплантации.

Рис. 6. Схема перехода от стадии двух клеток к стадии образования полости и затем к стадии обособления трофэктодермы и внутренней клеточной массы. Эмбрион заключен в плотную желеобразную «капсулу», прежде служившую оболочкой яйцеклетки; она носит название вителлинового слоя, или zona pellucida
Образ жизни эмбриона человека близок к паразитическому, но не стоит принимать эту метафору слишком близко к сердцу. В любом случае это единственный способ вынашивания потомства, а значит, выживания у нашего вида. Мать не просто терпит паразитизм ребенка, а поощряет его за счет постоянного взаимодействия. Если построенный на сигнальных молекулах «диалог» между маткой и эмбрионом прекращается, эмбрион не может оставаться имплантированным, и беременность прерывается.[24]
Иногда эмбрион движется по женским половым путям от места оплодотворения в маточной (фаллопиевой) трубе чересчур медленно и не достигает матки к тому времени, когда уже готов к имплантации. Одной из распространенных причин этого медленного перемещения является повреждение выстилки маточной трубы бактериями Chlamydia.[25] В наше время это часто встречается у молодых женщин.[26] Эмбрион, достигший определенной стадии развития, пытается имплантироваться там, где он находится в данный момент, даже в яйцеводе. Это ведет к внематочной беременности. Маточная труба не приспособлена для поддержания роста плода ни физически (по размеру и растяжимости), ни физиологически (в плане возможности обеспечить питание и приток крови). Во многих случаях это приводит к выкидышу. Иногда выкидыш приходится вызывать искусственно для спасения жизни матери.
До начала специализации клетки внутренней клеточной массы могут дать начало любой клетке организма. Во всяком случае, так происходит у мышей: на людях подобные эксперименты не могут проводиться по этическим соображениям. Для описания спектра типов клеток, в которые может дифференцироваться та или иная клетка эмбриона, исследователи часто используют специальную древовидную диаграмму, отображающую типы клеток и последовательность дифференцировки. Пример такой диаграммы приведен на рис. 7. Все возможные варианты в конечном счете происходят от одного типа клеток, который на такой диаграмме представляет собой ствол дерева. Поэтому эти клетки называют стволовыми клетками. (Это буквальный перевод немецкого термина «Stammzelle», который предложил в 1909 г. Александр Максимов, изучавший «древо» клеток крови.) В зависимости от конкретной схемы термин «стволовые клетки» может быть применен как к клетке, которая может создать только несколько различных типов клеток организма, так и к клеткам с большими потенциями.

Рис. 7. Типичная древовидная диаграмма, которая обобщает данные о траекториях клеточных дифференцировок – клетки ранних стадий развития дают начало клеткам более поздних стадий (или дифференцируются в них). Эта диаграмма, которую следует читать снизу вверх, отражает события, описанные в этой и последующих главах. Древо всего процесса развития человека было бы огромным и содержало бы сотни ветвей. Нижняя точка таких диаграмм всегда принимается за «стволовую клетку» независимо от того, находится ли эта клетка на самом раннем этапе развития или уже прошла определенное расстояние «вверх» вдоль ствола дерева. Об использовании термина «стволовые клетки» будет рассказано позже
В последнее время понятие «стволовые клетки» несколько сузилось. Многие авторы утверждают, что стволовыми клетками следует называть только клетки, способные поддерживать собственную популяцию, а также давать начало другим типам клеток, расположенным на диаграмме над ними. (Термин, предложенный Максимовым, в явном виде не предполагал подобной трактовки).[27] К счастью, эта терминологическая тонкость не влияет на применимость термина «стволовые клетки» к внутренней клеточной массе, которая показана на рис. 7 в нижнем правом углу, потому что ее клетки могут поддерживать свою популяцию, даже если удалить их из эмбриона и поместить в пробирку. Они дают начало всем тканям организма. Поэтому клетки внутренней клеточной массы иногда называют эмбриональными стволовыми клетками (ЭСК), особенно в тех случаях, когда их извлекают из эмбрионов и выращивают в культуре.[28]
Эмбриональные стволовые клетки сыграли очень важную роль в медико-биологических исследованиях последних десятилетий. В хорошо отработанных рутинных экспериментах ученые направленно «редактируют» гены ЭСК мышей, а затем вводят такие модифицированные клетки в клеточную массу эмбриона обыкновенной мыши. В результате в организме мыши, которой сделали инъекцию, образуется смесь нормальных клеток внутренней клеточной массы и генетически модифицированных ЭСК. Как правило, удается сделать так, чтобы часть сперматозоидов самца мыши развивалась именно из генетически модифицированных клеток. Потомство такого самца, полученное в результате обычного спаривания, будет иметь модифицированные гены во всех клетках тела. Этот метод позволил ученым создать мышиные «копии» генетических заболеваний человека, а эксперименты, проведенные на таких копиях, дают возможность лучше понять болезнь и разработать способы лечения.[29] Всего в лабораториях мира были получены десятки тысяч таких мышей. Многие факты, описанные в этой книге, были открыты в процессе наблюдения за протеканием беременности и эмбриональным развитием генетически модифицированных мышей.
Существуют и человеческие ЭСК.[30] Есть надежда, что, если найти способы управления их развитием и получения нужных типов клеток, их можно будет использовать для «ремонта» поврежденных тканей в организме человека или даже для создания новых тканей для пересадки. Тем не менее эти разработки воспринимаются неоднозначно: ЭСК можно получить, только уничтожив человеческий эмбрион на раннем этапе развития. Для некоторых людей тот факт, что эмбрион впоследствии становится человеком, является достаточным основанием для придания ему статуса человека. В рамках такого подхода уничтожение эмбриона, по сути, является убийством; оно неприемлемо, какую бы пользу ни сулило. Кто-то, напротив, считает, что у раннего эмбриона отсутствуют такие человеческие атрибуты, как мысли или чувства, и он не нуждается в защите. Тем не менее многие придерживаются промежуточной позиции и допускают возможность исследований на человеческих эмбрионах при условии, что это регулируется определенными правилами. Последние разработки могут помочь найти выход из этой этической дилеммы. Исследовательская группа из Японии недавно открыла способ превращать обычные соматические клетки мыши в клетки, подобные ЭСК. Они получили название индуцированных плюрипотентных стволовых клеток, сокращенно ИПСК.[31] Чтобы проверить, насколько они сходны с ЭСК, исследователи заменили на них внутреннюю клеточную массу эмбриона. В результате родились здоровые мышата. Этот метод относительно прост и в настоящее время используется во многих лабораториях по всему миру. Многие ученые применяли его к соматическим клеткам человека, создавая на их основе нечто очень похожее на ИПСК. Я говорю «нечто очень похожее», так как по этическим причинам нельзя экспериментировать с человеком, как с мышью, и точно проверить это нельзя. Если ИПСК действительно идентичны ЭСК человека, их можно будет использовать для того же, для чего сейчас используются ЭСК. Таким образом, отпадет необходимость использовать человеческие эмбрионы в исследованиях подобного рода. Это может воплотиться в жизнь, но вряд ли обрадует тех, кто присваивает эмбриону статус человека на том лишь основании, что эмбрион может им стать. Если все наши клетки могут стать плюрипотентными и дать начало человеческому организму, почему бы не считать людьми и их?
У нормального эмбриона клетки внутренней клеточной массы очень недолго остаются в исходном состоянии: уже через короткий промежуток времени они начинают специализироваться, и каждый из типов клеток приобретает свои отличительные черты. Наличие у эмбриона особой заполненной жидкостью полости означает, что некоторые клетки внутренней клеточной массы, которая лишь незначительно увеличивается в размерах, окружены другими, а некоторые имеют свободную поверхность. Наличие свободной поверхности снова нарушает симметрию межклеточной адгезии, так что клеточный слой, соприкасающийся с жидкостью, стоит особняком от других клеток. Он становится плотным слоем – гипобластом.[32] Раньше считалось, что клетки превращаются в гипобласт в силу того, что просто оказываются в нужное время в нужном месте. Однако недавние эксперименты на мышах показали, что некоторые клетки внутренней клеточной массы могут иметь к этому предрасположенность. Как только где-то возникает свободная поверхность, эти клетки мигрируют туда, а другие переходят внутрь, на их место.[33] В любом случае определяющим фактором пространственной организации служит именно свободная поверхность (рис. 8). Некоторые клетки гипобласта остаются там, где образовались, но большинство распространяются по слою трофэктодермы и формируют полый желточный мешок (это название используется по аналогии с эмбрионами низших позвоночных).
Оставшиеся клетки внутренней клеточной массы делятся на два слоя. Клетки, непосредственно прилегающие к гипобласту, остаются на месте и образуют новый слой – эпибласт. Клетки вышележащего слоя отслаиваются от эпибласта, и в результате формируется еще одна полость. Она называется амниотической полостью (см. рис. 8). Двухслойный диск, состоящий из эпибласта и гипобласта, зажат между полостью желточного мешка и амниотической полостью, как перемычка в греческой букве тета (θ). Эпибласт, слой, обращеннный к полости желточного мешка, даст начало всем клеткам будущего ребенка.[34],[35]

Рис. 8. Эмбрион снова проделывает «трюк со свободной поверхностью», чтобы получить из граничащего с жидкостью слоя клеток внутренней клеточной массы новый тип клеток – гипобласт. Слой оставшейся внутренней клеточной массы, соприкасающийся с гипобластом, снова дифференцируется, образуя эпибласт. Благодаря этому слой вышележащих клеток отделяется, образуя новую полость. Эпибласт – невзрачный клеточный диск – даст начало будущему младенцу; все остальное пойдет на образование тканей, необходимых для поддержания жизни плода в матке
События, описанные в этой главе, в основном вели к нарушению единообразия и появлению различий между ранее идентичными клетками. На ранней стадии развития эмбрион несколько раз использует один и тот же трюк – дальнейшая судьба клеток зависит от наличия или отсутствия у них свободной поверхности. Это позволяет использовать чистую геометрию в качестве источника новой информации. В каждом случае сугубо локальные взаимодействия приводят к масштабным изменениям. При этом ни одна клетка не «руководит процессом» и не «видит» его общей картины. С появлением разных типов тканей эмбриону будет значительно легче создавать дальнейшие различия. Например, новый тип клеток – С – может развиваться в местах контакта клеток типов А и B. Таким образом, сформируются две новые зоны контакта (АС и СВ), и каждая из них будет служить основой для спецификации следующих типов клеток. На более поздних стадиях развития «эпоху внешних воздействий» (например, использование свободных поверхностей) сменяют механизмы, основанные на внутренних различиях. Первый из них и, возможно, наиболее примечательный станет следующей темой для обсуждения.
Глава 4
Закладка плана строения
Самый важный момент в вашей жизни – не рождение, не свадьба или смерть, а гаструляция.
Самый важный момент в вашей жизни – не рождение, не свадьба или смерть, а гаструляция.
Что видят люди, оглядываясь на прожитые годы? Долгие периоды однообразной жизни, перемежаемые всплесками резких изменений. Возможно, эти изменения являются результатом долгой и тщательной подготовки, занимающей месяцы или годы, но эта подготовка скрыта от глаз. Лепет ребенка перерастает в нечто более осмысленное почти незаметно для родителей, но они никогда не забудут его первое слово. Постепенно складываются и отношения семейной пары: сначала возникает привязанность, затем растет доверие, возникает взаимопонимание, и, наконец, они осознают, как много значат друг для друга. Накопление профессиональных навыков гораздо менее очевидно, чем новая работа или продвижение по карьерной лестнице, которое мы получаем в награду. Гораздо менее приятный пример – постепенное накопление повреждений на клеточном уровне, незаметно подводящее к рубежу, когда можно установить точный диагноз и здоровый человек становится больным.
Такое чередование рутинных событий со скачкообразными изменениями свойственно не только жизни взрослого человека, но и жизни эмбриона. На ранних этапах развития эмбриона, описанных в предыдущих двух главах, происходило всего лишь простое деление клеток. Затем – на первый взгляд, внезапно – начались новые процессы, связанные с формированием клеточных слоев, заметно отличающихся друг от друга. Получившаяся в результате структура – по сути, наполненная жидкостью сфера, которая разделена на две камеры парой дисков (рис. 9), – по-прежнему далека от того, чтобы в ней можно было опознать что-то человекоподобное. Скажите кому-нибудь, что из этой пары дисков получится животное, и он, вероятно, предположит, что это будет медуза. По крайней мере, ее колокол, как и диски эмбриона, радиально симметричен, и у нее есть верх и низ, но нет оси, разделяющей тело на половинки (см. рис. 9). Тем не менее эмбрион уже успел подготовиться к масштабной реорганизации, после которой он уже будет напоминать человека. Этот процесс, занимающий всего пару дней, называется гаструляция.

Рис. 9. Простая радиальная симметрия зародышевого диска человеческого эмбриона в сравнении с аналогичной симметрией медузы
Чтобы подойти к теме гаструляции, можно было бы сначала рассмотреть строение примитивного организма, который образуется в результате данного процесса, а затем посмотреть, какой вклад в его образование вносит каждый этап гаструляции. Это, пожалуй, был бы самый рациональный и простой подход к описанию гаструляции, однако он может создать ложное впечатление о том, что клетки имеют какое-то представление об анатомии организма, в строительстве которого участвуют. На самом же деле развитие организма основано вовсе не на том, что клеткам известны законы развития (понять эти законы трудно даже нам, с нашим мозгом из миллионов клеток), а на том, что клетки автоматически откликаются на изменения окружающей их среды. Поэтому в этой главе мы, следуя логике развития, сначала рассмотрим поведение клеток развивающегося организма и только потом посмотрим, к чему оно приводит.
Прежде чем начать рассказ, хочу сделать важное предупреждение: изучать процесс гаструляции у человека крайне сложно, поэтому почти все, что описано в этой главе, основано на исследованиях животных. Существуют строгие правовые ограничения относительно выращивания человеческих эмбрионов в лабораторных условиях, и изучать эмбрионы на стадии гаструляции нельзя (почему – я расскажу чуть позже). Основная последовательность анатомических изменений человеческого эмбриона известна. Эти данные были получены в исследованиях немногочисленных (и очень ценных) эмбрионов, извлеченных при посмертном вскрытии или при удалении матки у женщин, которые чаще всего даже не знали о том, что беременны. Гаструляция начинается примерно через пятнадцать дней после оплодотворения, то есть примерно тогда, когда женщина ожидает начала менструации. Некоторым из этих эмбрионов более ста лет, но они до сих пор хранятся в музеях под особым присмотром, потому что новый материал такого рода поступает крайне редко. Исследования проводятся главным образом на мышах и курицах, а процесс гаструляции у этих животных значительно отличается от гаструляции у человека. Цыпленок развивается в яйце, а не в матке; эпибласт и гипобласт мыши имеют форму чаши, а не диска. Эти особенности могут играть существенную роль при развитии. Поэтому реконструировать механизмы гаструляции человека на основе данных, полученных при изучении модельных животных, рискованно – слишком мало у нас информации, и мы можем неверно интерпретировать какие-либо детали процесса.
Отправной точкой для гаструляции является эмбрион в том состоянии, в котором мы оставили его в конце главы 3. К этому времени уже сформировался ряд вспомогательных тканей, таких как плацента, а также два заполненных жидкостью внутренних пространства: амниотическая полость и желточный мешок. Между этими полостями лежат два диска, расположенные один над другим: гипобласт и эпибласт. Из гипобласта образуются дополнительные вспомогательные ткани, а из эпибласта – сам плод. Ни один из этих двух дисков не имеет каких-либо явных особенностей, которые позволяли бы отличить один его край от другого (см. рис. 9).
Первое изменение (судя по данным, полученным на животных) происходит в гипобласте. У клеток, расположенных в середине этого диска,[36] включаются новые гены, в том числе и ген ДНК-связывающего белка, который называется HEX.[37] Что является сигналом к этому изменению, пока неясно. Возможно, что все клетки гипобласта изначально готовы к нему, но большинство из них ингибируется сигнальным белком, который синтезируется тканями, окружающими гипобласт.[38] Достаточно далеко от источника этого ингибитора находятся только клетки в центре диска, поэтому они могут избежать его влияния и запустить экспрессию гена HEX. Этот механизм – только предположение, но активация гена HEX – факт. Клетки, в которых экспрессируется HEX, уходят из центра диска – они перемещаются, расталкивая соседние клетки, и собираются в одной точке на краю диска гипобласта[39],[40],[41] (рис. 10). Все еще непонятно, даже в случае мышей, что же особенного в этой точке диска, что клетки мигрируют именно туда. У «низших» животных расположение похожей «особой» точки обусловлено начальными условиями развития. В некоторых случаях положение этой точки определяется переходящей к зародышу асимметрией распределения молекул, заложенных в яйцеклетку материнским организмом. В других случаях в этой точке располагаются полярные тельца, «побочные продукты» клеточных делений, предшествующих формированию яйцеклетки. У некоторых организмов эту точку, по-видимому, маркирует место проникновения сперматозоида. Весьма вероятно, что у млекопитающих это тоже так, и есть указания на то, что эмбрионы мышей асимметричны уже на самых ранних стадиях развития.[42] Соответствующих данных по эмбрионам людей, разумеется, нет. Это очень досадно, потому что скопление в одном месте клеток, у которых экспрессируется HEX, приводит к исключительно важному результату – одно конкретное место на краю диска гипобласта становится не таким, как другие. Иными словами, это первый шаг в сторону от простой радиальной симметрии эмбриона (рис. 10).
Достигнув края диска гипобласта, группа клеток, экспрессирующих ген HEX, становится новой структурой ПВЭ.[43] Клетки ПВЭ начинают секретировать собственные сигнальные белки. Эти белки, способные распространяться на небольшие расстояния, вполне могут достигнуть эпибласта, лежащего непосредственно над гипобластом, явно в пределах дистанции распространения сигнала.[44] На этом этапе клетки эпибласта уже реагируют на сигналы, поступающие от вспомогательных тканей эмбриона. Эти сигналы подготавливают их к тому, чтобы создать структуры, характерные для задней части организма. Если бы больше ничего не происходило, весь эпибласт формировал бы структуры задней части, и это имело бы катастрофические последствия для эмбриона. Сигналы, поступающие от ПВЭ, уравновешивают эту тенденцию, запуская процессы создания тканей головы.[45] Если ПВЭ не будет синтезировать сигнальные белки, эмбрион не сможет правильно сформировать голову. Таким образом, положение ПВЭ в уникальной области на краю диска гипобласта диктует полярность вышележащего эпибласта: сторона, ближайшая к ПВЭ, сформирует голову, а удаленная от нее – заднюю часть тела эмбриона. Только небольшая часть эпибласта удалена от ПВЭ настолько, что вообще не получает от нее сигналов. По-видимому, это очень важно для локализации области, в которой происходят первые заметные изменения эпибласта. В этом месте клетки эпибласта переключаются на образование сигнального белка, который привлекает окружающие клетки – они мигрируют по направлению к источнику этого белка.[46] В результате такого перемещения клеток образуется важная структура со скромным названием – первичная полоска (см. рис. 10).

Рис. 10. Нарушение радиальной симметрии. Клетки в центре гипобласта «включают» ген HEX, движутся к краю и концентрируются в одном месте. Это нарушает радиальную симметрию гипобласта, а поскольку его клетки синтезируют сигнальные белки, которые «вмешиваются» в процессы, происходящие в эпибласте, его радиальная симметрия тоже нарушается
Особая роль ранней активности клеток ПВЭ гипобласта в определении места формирования первичной полоски была показана в экспериментах на куриных эмбрионах. После того как у них начинала нарушаться радиальная симметрия, гипобласт поворачивали относительно передне-задней оси эмбриона.[47] Эти манипуляции приводили к тому, что эпибласт формировал первичную полоску, подстраиваясь под новые координаты, а значит, именно гипобласт контролирует этот процесс.

Рис. 11. Удлинение первичной полоски и образование узелка (что такое узелок, я объясню далее в этой главе)
Так как все больше и больше клеток эпибласта мигрирует в область первичной полоски, они начинают расталкивать друг друга и встраиваться между соседями. Это приводит к постепенному удлинению первичной полоски от края диска внутрь по его радиусу (рис. 11). Поскольку в эту миграцию вовлекается все больше клеток, на краях диска их становится меньше. В результате, по мере того как полоска удлиняется, диск в целом начинает сужаться. Он приобретает овальную форму, и радиальная симметрия эпибласта теряется (см. рис. 11). В длинной оси овала угадывается будущая ось тела, которая проходит от макушки до последнего позвонка (если бы речь шла о животных, можно было бы сказать «до кончика хвоста», но применительно к человеку это будет звучать немного странно). Та часть первичной полоски, которая образуется первой (она находится на краю диска), становится задней частью тела, а часть, которая образуется последней (примерно в середине диска, теперь имеющего овальную форму), становится головой. Таким образом, формирование нашего тела начинается с седалища, а создание головы откладывается «на потом», и это не единственный случай, когда эмбриология напоминает нам о том, что не стоит воспринимать себя слишком серьезно.
Роль ПВЭ в определении места образования первичной полоски имеет одно интересное следствие. Если клетки, экспрессирующие ген HEX, образуют не одно, а два скопления на краю диска, то сформируются две отдельные небольшие ПВЭ, а значит, два центра распространения сигналов. Тогда возможно образование двух первичных полосок. Это третий – и самый редкий – тип близнецов, который встречается менее чем в 1 % случаев двуплодной беременности.
Если образуются две независимые первичные полоски, то они дадут начало двум полностью независимым осям тела, а значит, двум организмам. Такие близнецы, возникшие из одного и того же эпибласта после образования амниотической полости, будут иметь общую амниотическую, а также хорионическую полость. Этим они отличаются от близнецов, описанных в главе 2 (у них нет общих полостей), и от близнецов, описанных в главе 3 (они имеют общую хорионическую полость, но амниотическая полость у каждого из них своя). (Именно по числу общих полостей акушеры могут понять, с каким случаем возникновения близнецов они имеют дело.) Возникновение близнецов за счет двух первичных полосок в одном и том же эпибласте чревато опасностями, так как между ними нет четкой разграничительной линии и существует серьезный риск, что они не разделятся полностью. В этом случае на свет появятся сиамские, или сросшиеся, близнецы. Как правило, они не просто «срослись» друг с другом, но и имеют общие части тела. Общими могут оказаться в том числе жизненно важные внутренние органы. В принципе, сросшиеся близнецы могут быть вполне здоровыми людьми, хотя, конечно, их жизнь простой не назовешь. Пожалуй, самые известные сиамские близнецы – это Чанг и Энг Банкер[48],[49] (1811–1874 гг.), много лет гастролировавшие с цирком Барнума. В те времена глазеть на уродов считалось нормальным, и бородатые женщины, карлики, великаны и аномально тучные люди в цирках выступали на потеху публики. Братья Банкер назвали свой «дуэт» в честь родной страны, и за феноменом сросшихся близнецов с тех пор закрепилось название «сиамские близнецы». Родись братья в наши дни, их, скорее всего, разделили бы, но во многих других случаях разделить сросшихся близнецов очень трудно или вообще невозможно. Иногда это возможно только ценой жизни одного из близнецов. Поэтому операции по разделению сросшихся близнецов, помимо чисто технических проблем, нередко осложняются серьезными этическими вопросами.
Еще более сложные ситуации возникают в тех случаях, когда эпибласт образует не две отдельные первичные полоски, которые дают начало двум осям тела, а одну «недоудвоившуюся» Y-образную первичную полоску. Это приводит к образованию двух голов и, возможно, двух шей при общем позвоночнике. Это явление называется «удвоением оси», но название явно неудачно, так как удваивается не вся ось (в этом случае проблем бы не было), а только ее часть. У людей это явление встречается редко – как правило, оно приводит к выкидышу или мертворожденному ребенку, а если младенец рождается живым, то скоро умирает. Заспиртованные экспонаты такого рода есть почти во всех старых коллекциях естественно-научных редкостей. Очень редко люди с этой аномалией все же выживают. Самый яркий пример – это сестры Абигейл и Бриттани Хенсел, которым сейчас около двадцати пяти лет. Голова и шея у каждой из сестер своя, а туловище общее. Все, для чего нужна голова, они могут делать совершенно независимо, например, каждая из сестер может читать независимо от другой. При ходьбе, игре на пианино и управлении автомобилем им приходится действовать вместе (надо полагать, чиновникам пришлось немало потрудиться над разработкой индивидуального экзамена по вождению). Несмотря на анатомические аномалии, сестры Хенсел абсолютно нормальные, умные и общительные молодые женщины. Удвоение оси более широко распространено у других животных, например рептилий и амфибий, которые выживают с этой аномалией гораздо чаще людей. Двуглавый крысиный полоз по имени Мы (We) прожил в Городском музее Сент-Луиса восемь лет и стал одной из его главных достопримечательностей. Известность ему принесли не только необычный внешний вид, но и «споры», время от времени возникавшие между его головами, каждая из которых имела свое мнение. Есть даже одна ископаемая находка взрослой двухглавой ящерицы.[50] Удвоение оси у амфибий можно индуцировать путем воздействия на сигнальные механизмы, эквивалентные сигнальным механизмам в эпибласте и гипобласте млекопитающих (собственно говоря, существование этих механизмов было впервые показано именно на лягушках). Например, в результате введения в эмбрион лягушки одного из белков, синтезируемого в ПВЭ млекопитающих и необходимого для формирования головы, получается лягушка с несколькими головами. Этот белок получил название Cerberus («Цербер») в честь многоголового пса, стража входа в Аид.
С точки зрения этики закономерность «одна первичная полоска – один человек» может быть истолкована как доказательство того, что ее образование – поворотный момент в развитии человека. Главным аргументом является то, что до формирования первичной полоски нельзя точно сказать, сколько людей получится из эмбриона, а значит, установить четкую связь между конкретным эмбрионом и конкретным человеком. Развивая эту мысль, можно прийти к выводу, что структура, которую нельзя приравнять к конкретному человеку, не может быть наделена правами человека, а значит, проводить исследования эмбрионов до наступления стадии гаструляции можно. Напротив, как только сформировалась первичная полоска, количество будущих детей известно. На них распространяются права человека, и экспериментировать на них нельзя. Рассуждений такого рода немало, и некоторые из них сдвигают границу допустимого на более ранний период, вплоть до момента зачатия. Основная проблема такого подхода в том, что законодатели хотели бы видеть четкую границу между стадией, когда эмбрион еще не является человеком, и стадией, когда он бесспорно им уже является. Некоторые аспекты развития действительно связаны со ступенчатыми изменениями, как я уже говорил в начале этой главы, а другие (например, размер) меняются постепенно. Вполне возможно, что становление индивидуальности не является ступенчатым. Не исключено, что потенциальная способность стать человеком воплощается в реальную человеческую природу за счет последовательности шагов, занимающей месяцы и даже годы (формирование нейронных связей в мозге продолжается в течение долгого времени после рождения). Суть проблемы в том, что мы пока что слишком мало знаем о биологических основах человеческой индивидуальности, чтобы с уверенностью сказать, является ли ее становление внезапным или постепенным процессом. Вот почему в настоящее время все этические споры, которые якобы должны четко указать, когда именно человек становится человеком, колеблются на грани софистики.
Движения, благодаря которым клетки собираются вместе для формирования первичной полоски, не прекращаются и после ее создания. По мере поступления большего количества клеток центр полоски как бы проседает. Из полоски получается узкая бороздка, которая ближе к середине диска расширяется, образуя уплощенную область, называемую узелком. Клетки узелка продуцируют сигнальные белки, которые, во-первых, привлекают в этот регион дополнительные клетки, а во-вторых, заставляют их ослабить межклеточную адгезию и «включить» гены, способствующие их превращению в клетки других типов. Клетки, расположенные ближе к источнику сигнала (то есть к узелку), получают большую дозу сигнальных белков и реагируют сильнее. Ослабление системы адгезионных связей между клетками и активация у них миграционного поведения приводят к тому, что клетки свободно перемещаются в плоскости эпителиального диска и могут даже покинуть его.[51],[52],[53] Клетки проваливаются под поверхность диска эпибласта, переходя на его нижнюю сторону. Этот процесс распространяется по первичной полоске в направлении от головы к хвосту, так что более задние участки тела проходят его значительно позже, чем передние.[54] По мере того как ближайшие к центру бороздки клетки уходят из диска, их бывшие соседи сдвигаются ближе друг к другу и к центру бороздки так, чтобы заполнить свободное пространство, а затем, когда наступает их очередь, тоже проваливаются под диск, покидая эпибласт.
Первые клетки первичной полоски, которые уходят из диска на определенном уровне оси «голова – хвост», присоединяются к гипобласту: они расталкивают его клетки и встраиваются между ними, так что клеточный состав центральной области этого слоя полностью изменяется (рис. 12). Оказавшиеся в этой области клетки образуют слой, который называется эндодерма. (Это слово в переводе означает «внутренняя кожа»; название связано с тем, что в дальнейшем этот слой образует трубку кишки и трубки ассоциированных с ним органов – печени и поджелудочной железы.) Клетки, которые уходят из полоски немного позже, слабо связаны друг с другом и не образуют непрерывный слой. Они образуют мезодерму (в переводе – «средняя кожа»), которая лежит между эндодермой и внешним слоем. Клетки первичной полоски, которые не успевают «утонуть» и после завершения гаструляции остаются в эпибласте, формируют эктодерму (в переводе – «внешняя кожа»).[55] Таким образом, первичная полоска и узелок не только показывают нам ось «голова – хвост» у эмбрионов, но и превращают один слой клеток в три разных слоя: эктодерму, мезодерму и эндодерму (см. рис. 12). Это основные слои тела почти у всех животных.[56]
Для того чтобы уйти из диска, клетки должны сначала добраться до первичной полоски, и поэтому время их погружения тесно связано с исходным расстоянием до нее. Клетки, располагавшиеся ближе к средней линии диска, быстро проходят короткое расстояние и выселяются из полоски, тогда как клетки, располагавшиеся вблизи краев диска, начинают выселяться значительно позже, когда первые клетки уже давно погрузились под диск. Время и пространство так тесно сплелись в процессе гаструляции, что трудно определить, какой из этих факторов играет более важную роль в судьбе клеток. Определяется их судьба еще до начала миграции за счет ранних сигналов от ПВЭ или она становится известной только после того, как клетки выселяются из диска и оказываются внутри эмбриона? По-видимому, судьба некоторых клеток предопределена еще до начала движения.[57],[58],[59],[60],[61] Примером может служить обсуждавшееся выше программирование области, в которой будет формироваться голова, с помощью сигналов от ПВЭ, или программирование клеток, удаленных от ПВЭ, на формирование первичной полоски. Насколько четко определена на этих ранних этапах дальнейшая судьба других клеток, непонятно. Дело усложняется тем, что в некоторых случаях «запрограммированная» судьба клеток является всего лишь наиболее вероятным путем их развития. Если в эксперименте поместить их в другую среду и подвергнуть воздействию других сигналов (что вполне возможно и в естественных условиях, если во время гаструляции они затеряются среди других клеток), такие клетки могут «передумать» и пойти совсем по другому пути развития. Эта область эмбриологии все еще полна нерешенных вопросов, которые ждут своих исследователей.

Рис. 12. Образование трех слоев тела – эктодермы, мезодермы и эндодермы – во время гаструляции. В верхней левой части изображен эпибласт эмбриона, у которого уже образовался узелок, как он виден со стороны амниотической полости. Гипобласт лежит под эпибластом и не виден. На основном рисунке изображена первичная полоска в разрезе (он обозначен пунктирной линией на рисунке в верхней левой части). Клетки эпибласта активно мигрируют в плоскости пласта, и траектории их движения сходятся в бороздке, получившейся из первичной полоски. Затем эти клетки выселяются из эпибласта и уходят в пространство между дисками. Часть из них (те, которые встраиваются между клетками гипобласта, расталкивая их от средней линии к краям) дадут начало эндодерме, а часть – мезодерме (новому слою клеток посередине между эпибластом и гипобластом). Клетки, не покинувшие эпибласт, формируют эктодерму
Почти сразу после образования эндодермы клетки, расположенные вдоль центральной линии диска гипобласта (то есть под первичной полоской), начинают двигаться вверх (рис. 13). Это клетки, которые выселялись из эпибласта непосредственно через узелок, с той его стороны, которая обращена к голове, а значит, это клетки, получившие самую высокую дозу сигнальных молекул узелка. Благодаря им эти клетки «запрограммированы» на то, чтобы покинуть эндодерму.[62],[63],[64] Сразу после этого они выстраиваются вдоль оси эмбриона и образуют плотный стержень – нотохорд.[65] Это одна из важнейших структур на ранних стадиях развития эмбриона. Чтобы понять, почему эта структура возникает так рано (а также то, почему она возникает вообще), нужно обратиться к эволюции животного мира.
Зоологи классифицируют животных, используя иерархическую систему. Основной единицей этой системы является вид (например, Homo sapiens/Человек разумный). Схожие виды образуют род (например, Homo). Похожие рода сгруппированы в семейства (например, Hominidae), семейства – в отряды (например, Приматы). Отряды, в свою очередь, объединяются в классы (например, Mammalia/Млекопитающие), классы – в подтипы (например, Vertebrata/Позвоночные), а подтипы – в типы. Самые первые классификации основывалась на простом сходстве, но, после того как благодаря Дарвину и Уоллесу прояснилась роль изменчивости и естественного отбора в возникновении кластеров схожих таксонов, классификации стали отражать эволюционные взаимоотношения. Тип Chordata (Хордовые), к которому относятся все позвоночные, включает всех животных, имеющих нотохорд на каком-то этапе развития. С точки зрения количества видов в типе хордовых преобладают позвоночные, но есть в нем и несколько беспозвоночных, оставшихся с раннего кембрия. Большинство из них довольно редкие организмы, но ланцетник встречается достаточно часто, и в некоторых районах Азии его даже употребляют в пищу. Ланцетники – животные примерно пяти сантиметров в длину, внешне похожие на рыб. Они не имеют костного скелета, но нотохорд сохраняется у них на протяжении всей жизни, укрепляя тело и обеспечивая опору для мышц.

Рис. 13. Образование нотохорда из клеток, расположенных вдоль средней линии эндодермы (напомним, что эндодерма образовалась из погрузившихся клеток первичной полоски)
Нотохорд, однако, не просто придает телу жесткость. Он состоит из особых клеток, выделяющих специальные белки. Поэтому нотохорд может служить источником сигналов при разметке плана строения эмбриона, а его расположение вдоль средней линии делает его идеально приспособленным для этой цели. Хордовые, включая позвоночных, активно используют эти сигналы для разметки внутренних тканей. Так, например, детерминируются различные типы нервных клеток в спинном мозге или соединительных тканей и мышц по обеим сторонам тела. Сигналы нотохорда настолько важны, что событиям, в которых они играют ключевую роль, посвящены несколько последующих глав (5, 7 и 9). В ходе эволюции позвоночных план строения хордовых значительно усложнился, но все эти усложнения, в конечном счете, основаны на способности клеток на ранних этапах развития получать и интерпретировать сигналы нотохорда. Поэтому нам уже никуда не деться от этой структуры. У взрослого организма нотохорд заменяется более сложной конструкцией, позвоночным столбом, но на раннем этапе развития мы не можем обойтись без его сигналов. Нотохорд – эмбриологическая «живая окаменелость» – утратил свою изначальную функцию механической поддержки, но остается важным элементом нашего развития. Он существует ровно столько времени, сколько нужно для того, чтобы он успел выполнить свою функцию. Позже нотохорд разрушается, а его остатки идут на образование межпозвонковых дисков, смягчающих нагрузку на позвоночник[66] (повреждение этих дисков приводит к болезненной межпозвоночной грыже).
Мало того что система «узелок/первичная полоска» определяет положение главной оси тела и образует первые слои тканей, она также выполняет еще одну важную функцию – нарушение зеркальной симметрии левой и правой половины тела.[67] Она делает это, перемещая жидкость в полости эмбриона очень неэффективным, но крайне полезным образом. Многие клетки животных обладают ресничками – маленькими, гибкими выростами. Каждая ресничка оснащена крошечным моторчиком, состоящим из белков, которые получают энергию из химических реакций и используют ее для воздействия на другие белки. Благодаря этим моторчикам реснички движутся – бьются. У одноклеточных животных биение ресничек позволяет им перемещаться в жидкости. В организме человека некоторые неподвижные клетки, напротив, используют реснички для перемещения жидкости. Например, реснички клеток дыхательных путей очищают легкие от слизи, а реснички клеток, выстилающих яйцевод, продвигают яйцеклетки и ранние эмбрионы к матке. У клеток узелка тоже есть реснички, они находятся на нижней стороне узелка и обращены в заполненную жидкостью полость.
Реснички клеток узелка имеют две необычные особенности. Во-первых, они отходят от клетки под углом примерно 45 градусов и направлены вниз и назад. Наклон назад обусловлен тем, что клетки «чувствуют» передне-заднюю полярность эмбриона.[68] Во-вторых, они бьются не так, как обычные реснички (их биение напоминает удар кнута), а совершают круговые движения, напоминающие раскручивание лассо перед броском. Эти вращения очень быстрые, около шестисот оборотов в минуту (что сопоставимо с числом оборотов двигателя автомобиля на холостом ходу). Ресничка всегда вращается по часовой стрелке (если смотреть от ее кончика в сторону клетки). Это связано с тем, что двигательные белковые комплексы обладают зеркальной асимметрией и могут присоединяться к ресничкам и вращать их только одним образом.[69] Расположение каждой реснички таково, что, когда она находится в нижней части описываемой ей окружности, она движется к левой стороне эмбриона, а когда она описывает верхний полукруг, то движется к его правой стороне (рис. 14). Пока что все симметрично, но близость реснички к клетке в верхней части описываемой ею окружности при движении вправо означает, что движение жидкости существенно замедлено силой вязкого сопротивления у поверхности клетки. Когда реснички находятся в нижней части окружности, далеко от поверхности клетки, эта сила минимальна и жидкость продвигается более эффективно. Так возникает дисбаланс между количеством жидкости, выталкиваемым вправо, и количеством жидкости, выталкиваемым влево. Это похоже на эффект создания боковой силы гребным винтом, хорошо знакомый владельцам одномоторных лодок. Ограниченное пространство между винтом и корпусом приводит к тому, что при включенном моторе лодка не только движется вперед, но и смещается в сторону (рис. 14). В переполненной гавани это нередко приводит к интересным последствиям.
Неравномерный поток жидкости, обусловленный характером биения ресничек, приводит к тому, что левая сторона узелка постоянно омывается свежей жидкостью из запаса в пространстве под ресничками.[70] Клетки могут получать из нее много микроэлементов (например, кальций). Правая же сторона узелка получает отработанную жидкость, с меньшим количеством полезных веществ. Белки, секретируемые клетками узелка, также будут уходить влево.

Рис. 14. Реснички, вращающиеся у поверхности, направляют поток жидкости влево. В верхней части рисунка представлен вид на узелок. Реснички направлены вниз под углом 45 градусов по отношению к будущей брюшной стороне эмбриона и назад к хвостовому концу тела. Они вращаются по часовой стрелке, описывая конус. В нижнем левом углу рисунка – вид с хвостового конца эмбриона. Положение вращающихся ресничек создает дисбаланс потока жидкости – она более эффективно перегоняется влево. В нижнем правом углу представлен вид со стороны кормы лодки и изображен аналогичный эффект, знакомый тем, кому приходилось управлять лодкой в ограниченном пространстве
Клетки узелка выделяют в жидкость, которая находится под ними, различные белки, в том числе Nodal – мощную сигнальную молекулу названную так по месту образования (от англ. node узелок). Nodal синтезируется в обеих частях узелка, но как только он выходит наружу поток жидкости тут же относит его влево. Кальций стимулирует образование этого белка, поэтому свежая, богатая кальцием жидкость, поступающая в левую часть узелка, способствуют образованию там большего количества белка Nodal. Он влияет на производство других белков, в том числе тех, которые влияют на экспрессию генов. Накопление белка Nodal на левой стороне эмбриона приводит к тому, что эта сторона активирует немного иной набор генов, чем правая сторона. Так нарушается зеркальная симметрия эмбриона.
Потеря идеальной зеркальной симметрии полезна для процесса создания человеческого организма. Если смотреть снаружи, то кажется, что правая половина нашего тела зеркально симметрична левой, но многие внутренние структуры расположены асимметрично. Асимметричны сердце и круги кровообращения, селезенка и поджелудочная железа смещены влево, а печень и аппендикс – вправо. Существует огромное количество мелких различий между левым и правым полушарием мозга. Некоторые признаки асимметрии видны невооруженным глазом. У мужчин, например, одно яичко (в большинстве случаев левое) ниже, чем другое.[71] Возможно, способность к асимметричному развитию и не была жизненно необходима для эволюции высших позвоночных, но почти наверное упростила ее. В противном случае нам, вероятно, потребовались бы парные органы (кроме, разве что, пищеварительного тракта, центральной нервной системы, пениса, влагалища и мочевого пузыря, а они и так расположены по срединной линии). Это привело бы к проблемам с размещением органов в теле. У рыб, с их удлиненным узким телом, пары органов могли бы расположиться друг за другом, но наземным или летающим животным это, скорее всего, было бы невыгодно.
Самое примечательное в этом механизме нарушения симметрии то, что асимметрия на уровне организма обусловлена, по большому счету, асимметрией на молекулярном уровне, а именно асимметрией белковых комплексов, обеспечивающих биение ресничек. Это один из немногих случаев, когда конкретное свойство молекулы переносится на весь организм. Этот механизм необычен, но его существование подтверждено убедительными доказательствами. Во-первых, вращение ресничек клеток эмбриона можно наблюдать непосредственно. Во-вторых, создаваемый ресничками поток можно смоделировать – сначала математически, а потом и практически, в экспериментах с искусственными ресничками. Поток жидкости был неоднократно визуализован – как в эмбрионах, так и в искусственных моделях – путем добавления окрашенных частиц. Немало исследований посвящено образованию и накоплению белка Nodal. Идею о существовании такого механизма подкрепляют и данные по мутациям. Мутации, нарушающие образование или вращение ресничек, приводят к появлению организмов с нарушенной асимметрией. У одной мутантной мыши (inv) реснички направлены в другую сторону (под углом 45 градусов не в сторону хвоста, а в сторону головы). Поток жидкости движется в основном вправо, и можно предположить, что строение тела мышей с этой мутацией будет всегда зеркально отличаться от нормального. Именно это мы и наблюдаем. С аналогичной инверсией рождаются и некоторые люди – судя по всему, по той же причине.
События, описанные в этой главе, происходят всего за два-три дня (спустя 15–17 дней после зачатия), но кардинально меняют строение эмбриона. Прежде он представлял собой диск, на вид не имеющий ничего общего со сложным животным. Теперь же у него есть удлиненное тело с четкой выраженным передним и задним концом, спинной и брюшной стороной и правым и левым боком, ткани трех типов, определенным образом расположенных относительно друг друга, а также нотохорд, проходящий по центральной оси. Задатки будущего животного налицо, и можно приступать к усложнению внутреннего строения.
Глава 5
Формирование мозга
Мозг – это орган, с помощью которого мы думаем, что мы думаем.
Мозг – это орган, с помощью которого мы думаем, что мы думаем.
Нервная система взрослого человека – невероятно сложная структура, отростки которой подходят практически ко всем остальным частям тела. Ее центральной частью является спинной мозг – длинная трубка диаметром около полутора сантиметров, проходящая по срединной линии спины (рис. 15). В голове основная структура спинного мозга изменяется и образует ряд вздутий, которые вместе формируют головной мозг. От спинного мозга отходят нервы, которые посылают сигналы мышцам и получают информацию от органов чувств. Кроме того, есть квазинезависимые части нервной системы, которые регулируют работу внутренних органов, таких как пищеварительная система и сердце. Однако спинной мозг в некоторой мере контролирует даже их.
Нервная система очень важна для функционирования взрослого организма. Во многих странах прекращение ее работы («гибель мозга») юридически считается смертью человека, даже если другие системы организма все еще поддерживаются искусственным путем. У эмбриона, однако, нервная система не играет заметной роли в контроле жизнедеятельности до самых поздних стадий развития. Тем не менее она начинает развиваться раньше, чем большинство других внутренних органов. Есть несколько возможных объяснений того, почему это так. Может быть, дело в том, что основные элементы архитектуры нервной системы, которая охватывает большую часть тела, должны быть заложены прежде, чем в тех же регионах сформируются другие структуры и органы. С другой стороны, раннее развитие нервной системы может быть связано с тем, что это одна из старейших частей тела с точки зрения эволюции.

Рис. 15. Рисунок, созданный Андреасом Везалием, анатомом эпохи Возрождения, и показывающий нервные пути, отходящие от спинного мозга к остальным частям организма. На рисунке не показан головной мозг, который в основном не связан напрямую с другими частями организма. Сигналы от головного мозга к органам тела и обратно проходят по спинному мозгу. (Головной мозг напрямую связан с сетчаткой и органами обоняния, но они, если посмотреть с точки зрения развития, и так по сути являются его частью.) Te Granger Collection/Topfoto
Как общее правило (с целым рядом исключений), порядок появления новых структур у эмбриона соответствует порядку, в котором они появлялись в процессе эволюции. Никто не знает наверняка, почему это так. Согласно одной популярной теории, дело в том, что вмешательство в основные механизмы развития на стадии формирования общего плана строения с большей вероятностью приведет к катастрофическим последствиям, чем вмешательство на стадии детализации уже существующих структур. Случайные мутации, которые влияют на основной план строения организма, скорее всего, приведут к его нежизнеспособности, а мутации, затрагивающие более поздние стадии, могут привести к появлению жизнеспособных организмов. Некоторые из них, возможно, смогут занять новую экологическую нишу и дать начало новому виду. В ходе эволюции новые виды возникали прежде всего за счет изменений на поздних этапах развития, и эмбрионы представителей разных видов гораздо больше похожи друг на друга, чем взрослые особи. Интересно, что эмбрионы на самых ранних стадиях развития (тех, о которых шла речь в главах 2 и 3) более восприимчивы к эволюционным изменениям. Возможно, это связано с тем, что изменения, происходящие еще до закладки плана строения, в меньшей мере чреваты катастрофическими последствиями. В конце книги приведен список дополнительной литературы на эту тему.
Как бы то ни было, развитие нервной системы начинается сразу после гаструляции. Точнее говоря, в передней (головной) части эмбриона она начинает формироваться уже тогда, когда в задних отделах продолжается гаструляция. Вся нервная система образуется из эктодермы эмбриона, а конкретнее, из полоски эктодермы, проходящей вдоль центральной линии спины от головы к хвосту. Клеткам этой полоски предстоит перестать быть эктодермой («внешней кожей») в строгом смысле слова и образовать внутреннюю нервную трубку, из которой потом получатся спинной и головной мозг. Более того, этот переход должен произойти без нарушения целостности эмбриона. Это яркий пример проблемы, упоминавшейся в главе 1: эмбриону нужно претерпеть существенные структурные перестройки и при этом не развалиться на части. Эта проблема решается за счет того, что некоторые клетки меняются местами с соседями, растягиваются и меняют форму, благодаря чему весь слой ткани складывается, как оригами.
Первый признак формирования нервной системы – заметное изменение формы эмбриона. Гаструляция уже нарушила простую форму диска, удлинив его по будущей передне-задней оси тела. После гаструляции удлинение продолжается: короткий и «коренастый» эмбрион становится вытянутым и тонким по мере того, как клетки, меняясь местами с соседями, перебираются все ближе к центральной линии (линии голова – хвост) и выстраиваются вдоль нее. На рис. 16, представляющем схему аналогичного процесса у плодовых мушек, показан один из способов того, как клетки, меняясь местами, могут изменить форму всей ткани.[72],[73] Клетки, которые раньше были отделены друг от друга другими клетками, например L1 и R1, становятся соседями, а клетки, которые жили бок о бок, например M1, M2 и М3, расстаются и расходятся в разные стороны. За счет этого процесса короткий и широкий участок ткани становится длинным и тонким.

Рис. 16. Изменение формы участка ткани за счет смены клетками соседей
Вероятно, клетки могут не только меняться местами, но и активно мигрировать в направлении центральной линии. Оба процесса осуществляются благодаря способности клеток воспринимать сигналы (большинство из них еще только предстоит расшифровать, хотя кое-какие мы уже знаем[74]), которые указывают им направление «голова – хвост» и «лево – право». Этот внутренний «компас»[75] позволяет клеткам ориентироваться в плоскости клеточного пласта. Благодаря работе этого компаса происходит так называемое конвергентное вытяжение эмбриона вдоль оси «голова – хвост»: клетки левой и правой сторон эмбриона конвергентно перемещаются к его центральной линии и выстраиваются вдоль нее. В результате тело эмбриона приобретает удлиненную форму, характерную для взрослого организма. В частности, широкий и короткий участок, расположенный вдоль центральной линии спины, становится длинным и тонким. В том месте, где образуется голова, он, правда, остается чуть более широким, так что тело эмбриона слегка напоминает по форме замочную скважину.
После стабилизации общей формы тела начинается активное развитие нервной системы. Нотохорд, который уже проходит вдоль центральной линии тела (глава 4), секретирует сигнальные белки. Эктодерма в радиусе действия этих сигналов (на практике это означает полоску эктодермы, лежащую непосредственно над нотохордом) готовится к превращению в нервную ткань. В клетках этой полоски включаются ранее неактивные гены; клетки немного утолщаются, готовясь к последующим изменениям формы. Образуется нервная пластинка. Ее клетки немного отличаются друг от друга в зависимости от положения: клетки, находящиеся непосредственно над нотохордом, образуют центральную полоску, а те, что находятся на границе с обыкновенной эктодермой, образуют две краевые полоски. На этом этапе полоски не отличаются внешне, но в дальнейшем они дадут начало соответственно центральной части и краям глубокой борозды (рис. 17).
Клетки нервной пластинки связаны друг с другом тем же типом межклеточных контактов, что и клетки эмбриона на ранней стадии развития, когда они образовывали компактную клеточную массу (глава 3). Внутри каждой клетки эти контакты соединяются друг с другом белковыми микрофиламентами, образующими непрерывную механическую сеть (рис. 18). Важно отметить, что эта сеть в основном залегает не в середине клеточного пласта, а вблизи обращенной наружу поверхности клеток.
Наличие этой сети означает, что никакая клетка не может изменить форму, не оказав при этом воздействия на окружающих ее соседей. Это, в свою очередь, означает, что действия отдельных клеток могут привести к изгибанию эпителиального пласта. Клетки центральной полоски производят большое количество белка Shroom, который взаимодействует с системой микрофиламентов, перестраивая ее так, чтобы межклеточные контакты подтягивались ближе друг к другу.[76] Если смотреть сбоку, профиль клеток центральной полоски изменяется от прямоугольного до клиновидного (рис. 19, а—б). Однако клетки по-прежнему крепко соединены межклеточными контактами, а значит, когда отдельные клетки приобретают клиновидную форму, промежутков между ними не образуется. Это приводит к скручиванию всего клеточного пласта[77] (см. рис. 19, б). В результате этого скручивания центральная линия эктодермы, расположенная вдоль спинной стороны эмбриона, прогибается внутрь, и образуется глубокая бороздка (рис. 19, в). Клетки краевых полосок, напротив, расширяют свои апикальные стороны и сужают базальные, (механизм этого процесса пока непонятен). Такое изменение формы клеток тоже приводит к изгибанию пласта, но уже в другую сторону – вместо борозды образуется валик. Оба процесса приводят к одному и тому же результату: область эктодермы, которая пойдет на формирование нервной ткани, прогибается внутрь и погружается вглубь эмбриона, а края этой области сдвигаются ближе друг к другу.

Рис. 17. Три полоски эктодермы, образующиеся на дорсальной (спинной) поверхности эмбриона, дают начало глубокой борозде

Рис. 18. Система микрофиламентов и межклеточных контактов образует единую механическую сеть, которая проходит через всю эктодерму, включая и нервную пластинку. Обратите внимание, что эта сеть находится ближе к апикальной (внешней) стороне клеток

Рис. 19. Этапы формирования нервной трубки. На всех рисунках показаны поперечные срезы эмбриона. Клетки с прямоугольным контуром (а) приобретают клиновидную форму, что приводит к образованию борозды (б—в), края которой в конечном итоге смыкаются, преобразуя борозду в трубку (г)
Образование центральной борозды – хороший пример масштабной перестройки тканей за счет локальных сил. Если вырезать из эмбриона кусочек дорсальной эктодермы, в которой и должен протекать этот процесс, и поместить его в чашку Петри, характерная борозда появится тогда, когда она и должна появиться у эмбриона. Это доказывает, что после того, как процесс запущен, он регулируется вовлеченными в него клетками, то есть локально, и контроль со стороны других частей эмбриона уже не нужен. На этом этапе, однако, развитие замедляется, потому что следующий этап зависит от активности эктодермы на краях центральной полоски, которая не будет сама по себе формировать нервную ткань. Ее клетки делают три важные вещи: они уплощаются, благодаря чему становятся короче, но шире; они размножаются; и они по-прежнему конвергируют (перемещаются) к срединной линии эмбриона, внося вклад в уже описанные процессы сужения и удлинения.[78] Все эти активности клеток способствуют созданию бокового давления, которое подталкивает края борозды друг к другу до тех пор, пока они не встретятся (рис. 19, в—г).
Когда это происходит, края двух сторон борозды смыкаются, и она превращается в трубку, пока еще соединенную с эктодермой. По мере смыкания краев борозды соседние области тоже сходятся, и борозда замыкается, как застежка-молния. Сразу после образования нервной трубки клетки перестраиваются так, что она полностью отделяется от окружающей ее эктодермы, не нарушая при этом целостности эмбриона. Подробности этого процесса до сих пор неизвестны, но, возможно, он происходит за счет механизма, основанного на клеточной адгезии. Он заключается в следующем. Адгезия между клетками нервной трубки, основанная на действии таких белков, как N-кадгерин, возможно, значительно сильнее, чем адгезия между этими клетками и клетками окружающей эктодермы. Поэтому клетки нервной трубки на границе с обычной эктодермой стараются установить как можно более прочный контакт с подобными себе клетками, а этого можно добиться только за счет ослабления контакта с окружающей эктодермой. После того как клетки эктодермы с противоположных сторон нервной трубки соприкоснулись, адгезия между ними, основанная на действии молекулы под названием Е-кадгерин, становится сильнее, чем адгезия между ними и клетками нервной трубки. Поэтому клетки эктодермы сильнее прилегают друг к другу, а контакт с клетками нервной трубки ослабляется. Такое объединение клеток по принципу «одного поля ягоды» не требует каких-либо особых механизмов, кроме простой биофизики клеточной адгезии. В конце концов две ткани просто теряют контакт друг с другом, и эктодерма спинной стороны эмбриона становится непрерывным слоем. Впоследствии он даст начало наружному слою кожи плода. Обращаю ваше внимание на то, что это гипотетический механизм. Пока что нет уверенности даже в том, что сила адгезионных взаимодействий между сходными клетками, вовлеченными в этот процесс, выше, чем между различными.
Смыкание нервной трубки у человеческого эмбриона требует филигранной точности. Нарушение этого процесса может привести к тому, что нервная трубка не сомкнется или сомкнется не полностью. Когда смыкания не происходит вдоль части спинного мозга, ребенок рождается с расщепленным позвоночником (spina bifida). В Шотландии, где была написана эта книга, этот порок развития в свое время был очень распространен и встречался у одного из ста детей, зачатых в определенных регионах. По крайней мере в четверти случаев основная проблема заключается в том, что две стороны нервной трубки не стыкуются, потому что боковая эктодерма не подталкивает их внутрь. Оказалось, что смыкание нервной трубки очень зависит от содержания фолиевой кислоты (витамин В9) в тканях. Если фолиевой кислоты мало, скорость размножения клеток низкая, и риск расщепления позвоночника высок. Название «фолиевая» означает «из листьев», и основными природными источниками этой кислоты являются зеленые овощи, бобы, некоторые плоды и семена. К сожалению, эти важнейшие продукты нередко практически отсутствуют в рационе жителей промышленных центров. Эта проблема стоит очень остро для малообеспеченных слоев городского населения Шотландии. Поэтому изучением влияния дополнительной фолиевой кислоты (в виде витаминных добавок) на будущих матерей занялось несколько исследовательских групп. Фолиевую кислоту давали, начиная со стадии планирования беременности и вплоть до окончательного формирования нервной трубки плода. Почти во всех опубликованных исследованиях результатом лечения стало резкое снижение риска возникновения расщепленного позвоночника (примерно на 50–75 %).[79],[80]
Общий вывод из этих исследований, а именно, что фолиевая кислота существенно сокращает риск серьезного порока развития, привел к тому, что всем женщин, планирующим беременность, стали рекомендовать принимать пищевые добавки с фолиевой кислотой. Однако этот простой совет бесполезен в случае незапланированной беременности, потому что смыкание нервной трубки происходит уже через три-четыре недели после зачатия, когда женщина, как правило, еще не знает, что она беременна. К сожалению, частота незапланированных беременностей и вероятность того, что беременность будет выявлена относительно поздно, наиболее высоки именно у беднейших и наименее образованных слоев населения, которые, как правило, к тому же едят очень мало свежих овощей. Поэтому в некоторых странах, включая США, основные продукты питания, например хлеб и зерновые завтраки, в настоящее время обогащают фолиевой кислотой, так что все население восполняет ее дефицит. Тем не менее требование государства добавлять в пищу какое-либо вещество, даже если речь идет о молекуле, которая встречается в природе и считается абсолютно безопасной, спорно с этической точки зрения. На момент написания этой книги ни в одной из стран Европейского Союза добавление фолиевой кислоты в продукты питания не являлось обязательным, но, например, сухие завтраки, обогащенные этим веществом, продаются повсеместно.
Как правило, расщепление позвоночника не смертельно, но, в зависимости от локализации и тяжести дефекта, ребенок может страдать от болезней разной степени тяжести; чаще всего возникает паралич нижних конечностей или недержание мочи. Неудачное смыкание нервной трубки в области головного мозга приводит к гораздо более серьезной болезни под названием анэнцефалия. При этом отсутствует затылок и большая часть мозга, а это уже смертельно. Как правило, ребенок с этим пороком развития рождается мертвым или умирает вскоре после рождения. Следует отметить, что недостаток фолиевой кислоты – не единственное, что может привести к расщеплению позвоночника и анэнцефалии. Важную роль играют также генетическая предрасположенность и влияние окружающей среды. Даже те матери, которые очень внимательно относятся к своему рациону и образу жизни, могут родить детей с этим пороком развития, и их вины в этом нет.[81]
Существует еще одна редкая, но примечательная аномалия, связанная с процессом смыкания нервной трубки. При некоторых типах возникновения близнецов два эмбриона могут развиваться очень близко друг к другу, но быть при этом разного размера. Очень редко случается так, что маленький близнец попадает в нервную трубку большого и оказывается навеки запечатан в ней.[82] Там его развитие может продолжиться – либо в виде неорганизованной опухоли, или в виде крошечного, но в принципе нормального плода. В одном из клинических отчетов описывается шестинедельный ребенок с необычным расширением головы. Внутри одной из нормальных полостей мозга ребенка находилось крошечное тело его близнеца с развитыми конечностями, туловищем и головой.[83] Это один из примеров явления fetus in fetu (буквально «плод внутри [другого] плода»). Несколько чаще (хотя все равно очень редко – примерно два случая на миллион рожденных младенцев) встречается другой тип этой аномалии: меньший близнец оказывается не в голове, а в животе большего, и происходит это на более поздней стадии развития. Такие случаи абдоминального fetus in fetu обычно диагностируются спустя несколько недель или месяцев после рождения, но могут оставаться незамеченными и более долгое время. Описан клинический случай,[84] когда человек вынашивал в животе своего близнеца в течение тридцати девяти лет.
Эти два нарушения формирования нервной трубки – расщепление позвоночника и крайне редкое явление fetus in fetu – имеют одну общую черту – они обусловлены, по крайней мере частично, факторами, не связанными непосредственно с эмбрионом. Расщепление позвоночника в той или иной степени связано с неправильным питанием матери, а явление fetus in fetu – с тем, что один из эмбрионов оказался именно там, где должна закрыться нервная трубка другого. Эти два примера подчеркивают тот важный момент, что эмбриональное развитие не является жестко детерминированным результатом генетической организации эмбриона. Оно обусловлено, даже на самых ранних стадиях, взаимодействием между генами и окружающей средой.
Нервная трубка, образованию которой была посвящена почти вся глава, еще не содержит ни собственно нервных клеток, ни нейронных связей. Нет еще ни чувств, ни рефлексов, ни разума, ни воли, ни мыслей – все это появится гораздо позже. Поэтому нервная трубка не может выполнять функции командования и управления, которые в один прекрасный день возьмут на себя головной и спинной мозг. Однако основной план их строения уже заложен, а кроме того, нервная трубка готова выделить клетки для построения нервной системы за пределами позвоночного столба. Это произойдет уже очень скоро, а как именно, мы узнаем в главе 7.
Общее удлинение тела, на фоне которого происходили описанные в этой главе события, играет важную роль в формировании другой трубки – кишки. Слой эндодермы, из которого образуется кишка, дифференцируется в процессе гаструляции (глава 4), но до удлинения эмбриона он представляет собой всего лишь плоскую пластинку на его вентральной (брюшной) поверхности, обращенную в просвет желточного мешка (рис. 20, а). Удлинение тела эмбриона растягивает эндодермальную пластинку, ее длина становится больше, чем ширина отверстия, ведущего в желточный мешок, а ее концевые участки подворачиваются с образованием коротких трубковидных отростков, направленных в сторону головы и хвоста эмбриона. По мере дальнейшего удлинения тела эти отростки тоже удлиняются, и в них начинают угадываться передняя кишка, из которой возникнет пищевод, желудок и верхние отделы кишечника, и задняя кишка, из которой образуются нижние отделы кишечника (рис. 20, б). Отверстие, ведущее в желточный мешок, казавшееся сразу после гаструляции огромным (относительно размеров эмбриона), сохраняет исходные размеры и поэтому вскоре принимает вид незначительного ответвления от удлиняющейся кишки растущего эмбриона (рис. 20, в). В конце концов это ответвление исчезает совсем, отверстие, ведущее в желточный мешок, закрывается, и кишка превращается в трубку, залегающую в глубине тела (а на ее концах образуются два новых отверстия – рот и анус).
После событий, описанных в этой главе, эмбрион уже обладает основными структурами хордового животного. У него есть удлиненное тело, три основных слоя тканей, нотохорд, дорсальная нервная трубка и вентральная трубка кишки (правда, еще не утратившая связи с желточным мешком). До конца развития еще очень далеко, но можно сказать, что начальный этап завершился. Менее чем за четыре недели одноклеточный эмбрион самоорганизовался в простое тело, состоящее из тысячи клеток. Совершенно самостоятельно он сперва создал различия там, где их не было, при помощи хитроумного использования простых геометрических законов, а затем использовал эти различия как источник информации для создания новых различий, новой информации, новых структур. Механизмы, за счет которых все это было достигнуто, относительно просты и подчиняются относительно простым правилам, действующим в локальном масштабе. Достигнув этой стадии умеренной сложности, эмбрион обладает достаточным количеством внутренней информации, чтобы в короткие сроки усложниться очень значительно. Его анатомическое строение сильно изменится, однако, как мы скоро убедимся, общий принцип развития не изменится вовсе: изменения структуры и дальше будут происходить за счет простых правил и локальных взаимодействий.

Рис. 20. Удлинение тела, сопровождающее формирование нервной трубки, важно также для образования кишечной трубки из исходно плоской эндодермальной пластинки. Удлиняющаяся спинная сторона эмбриона тянет за собой передний и задний края эндодермальной пластинки, в результате они изгибаются и заворачиваются в трубки. По мере дальнейшего роста эмбриона отверстие, ведущее в желточный мешок, становится все меньше и меньше, наконец превращается в незначительное ответвление кишечной трубки и закрывается. На стадии, изображенной на нижнем рисунке, кишка уже имеет различные ответвления, но они не показаны для облегчения восприятия
Глава 6
Долгое деление
Никакое дело не покажется невыполнимым, если разбить его на мелкие части.
Никакое дело не покажется невыполнимым, если разбить его на мелкие части.
Важной особенностью, о которой я еще не упоминал, является то, что регионы тела, расположенные на разных уровнях передне-задней оси, отличаются друг от друга. Некоторые структуры, например нервная трубка, простираются от головы до конца тела, но многие другие структуры появляются только на определенных уровнях этой оси. Если принять за целое расстояние от головы до хвоста, то руки, например, выступают из корпуса на уровне примерно три восьмых (у взрослых), а ноги растут фактически из конца этой оси. На участке туловища между руками и ногами конечностей нет. Органы внутри тела тоже располагаются строго определенным образом: глаза находятся на уровне одной восьмой расстояния от головы до хвоста, а печень – на уровне шести восьмых.[85] Все эти структуры развиваются из клеток эмбриона, и появление разных органов на разных уровнях оси «голова – хвост» говорит о том, что клетки эмбриона, расположенные на разных участках этой оси, отличаются между собой. В этой главе речь пойдет о том, как возникают эти различия. Чтобы понять механизмы этих процессов, нужно осознать одну важную, но малозаметную особенность человеческой анатомии.
Если приглядеться к беспозвоночным, таким как дождевые черви, многоножки или осы, то можно увидеть, что их тело состоит из серии повторяющихся сегментов. Тело червя, например, представляет собой длинный цилиндр со множеством поперечных линий, маркирующих границу между смежными сегментами (рис. 21, а). Большинство сегментов тела червя на вид одинаковы, хотя сегменты головы, хвоста и репродуктивной области все-таки немного отличаются от остальных. Многоножка похожа по форме тела на червя, но каждый сегмент ее тела несет пару ног. Головной и хвостовой сегменты многоножки тоже отличаются от других сегментов. Брюшко осы тоже сегментировано, что подчеркивают цветные полосы, а если посмотреть через лупу, то видно, что сегментирована и грудь. Она состоит из трех сегментов, которые отличаются друг от друга. От каждого из них отходит пара конечностей, но только два сегмента несут еще и пару крыльев (чтобы убедиться, что у осы четыре крыла, нужно внимательно присмотреться: задние крылья малы и находятся очень близко к задней кромке передней пары).

Рис. 21. Сегментация (а) дождевого червя, тело которого состоит из серии повторяющихся сегментов, и (б) человека, сегментация которого менее очевидна. Чтобы лучше продемонстрировать сегментированный характер позвоночника человека, на рисунке (б) не показаны конечности и ребра, а также голова и место соединения с ней (в том числе два самых верхних позвонка). Кроме того, изображение позвоночника немного стилизовано: на самом деле позвонки разных типов отличаются друг от друга рядом деталей
Деление тела на многочисленные сегменты, в основном схожие, но имеющие и определенные специализированные черты, зависящие от их положения в теле, является особенностью многих типов животных. У примитивных видов, например земляных червей, каждый сегмент удивительно самодостаточен с точки зрения физиологии: собственные простейшие легкие, почки, нервная система и т. д. Некоторые системы, например пищеварительная, проходят через все или большинство сегментов, а нервные системы разных сегментов соединены, и поэтому животное ведет себя как единое целое. Тем не менее формирование большей части тела за счет повторения базовых модулей дает большое преимущество, так как позволяет снова и снова использовать одни и те же генетические и клеточные системы. Благодаря такой экономии возникновение таких животных в процессе эволюции было менее маловероятным, чем возникновение животных, каждая часть которых уникальна. Сегменты наиболее схожи именно у самых примитивных животных (примитивных как в смысле простоты организации, так и с точки зрения раннего появления в палеонтологической летописи). В ходе дальнейшей эволюции появляется тенденция к специализации сегментов для выполнения особых задач, таких как полет и ходьба. По мере специализации сегменты становятся более взаимозависимыми с физиологической точки зрения, и на смену сегментам с собственной почкой, дыхательным аппаратом и т. д. приходят сегменты, полагающиеся на единые централизованные системы (выделительную, дыхательную и т. д.) всего организма. Позвоночные тоже сегментированы, хотя это не столь очевидно. Глядя на гладкое тело рыбы, трудно заподозрить, что она имеет сегментированное строение, но любой, кому случалось есть рыбу, замечал, что бо́льшая часть ее тела состоит из повторяющихся серий мышц, расположенных вдоль позвоночника, состоящего из повторяющихся серий позвонков и ребер. Люди, в чьей эволюционной родословной были похожие на рыб предки, тоже имеют сегменты, хотя их еще труднее разглядеть. Сегментированный характер строения человеческого тела проще всего увидеть на примере скелета, особенно позвоночника (рис. 21, б). Он состоит из тридцати трех позвонков, каждый из которых является вариацией на общую тему. Два верхних позвонка (в области шеи) служат для того, чтобы их владелец мог кивать и покачивать головой. В шейном отделе есть еще пять позвонков, далее идут двенадцать соединенных с ребрами позвонков в области груди. Еще ниже расположены пять массивных позвонков поясничного отдела, несущих значительную часть массы тела. И наконец, девять маленьких позвонков в самом низу позвоночного столба сливаются, образуя крестец и копчик. Мягкие ткани, которые крепятся к позвонкам, например мышцы, соединяющие соседние ребра, и нервы, идущие от разных частей тела к спинному мозгу, тоже имеют признаки сегментации. Сегментация прослеживается и в ряде других анатомических структур, но, как правило, на более тонком уровне.
В процессе эмбрионального развития сегментация человеческого тела начинается в мезодерме, лежащей слева и справа от нервной трубки. Клетки этой ткани, образовавшейся в процессе гаструляции (глава 4), вначале не очень прочно связаны между собой и не организованы каким-либо особым образом. В процессе сегментации, однако, эта ткань будет поделена на блоки, называемые сомитами, или первичными сегментами. Сомиты – предшественники позвонков, мышц и некоторых других структур тела. Они формируются не сразу, а последовательно, от шеи эмбриона к хвосту, и попарно (одновременно образуются два сомита – слева и справа от нервной трубки). У человеческого эмбриона каждая новая пара сомитов образуется примерно через шесть часов после предыдущей (рис. 22).
Задача превращения участка несегментированной ткани в сомит решается с помощью изменения адгезионных свойств клеток. Клетки, обладавшие высокими адгезионными свойствами в эктодерме до гаструляции, а затем превратившиеся в неплотную мезодерму, теперь повторно активируют свои адгезионные системы.[86] После этого они отделяются от своих бывших соседей по мезодерме и организуются в закрытые сферические структуры, похожие на цисты, – новые сомиты.
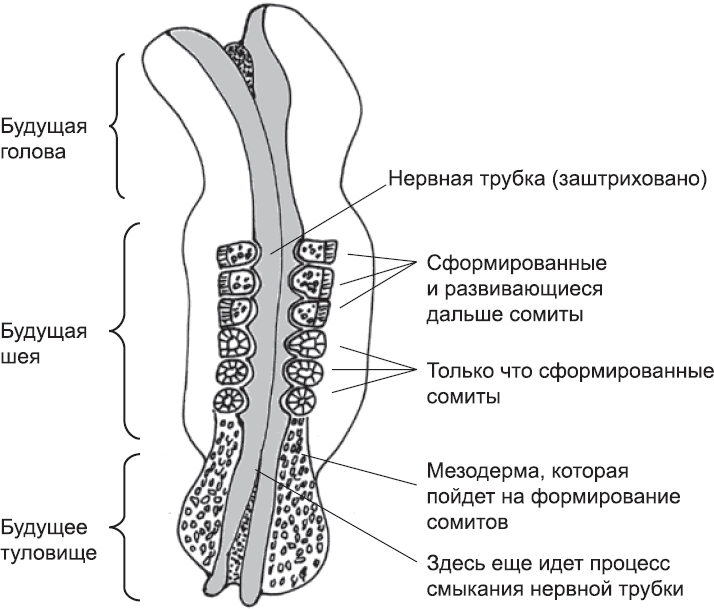
Рис. 22. Образование сомитов у человеческого эмбриона в возрасте примерно пяти недель. Уже сформированы шесть сегментов, которые дадут начало черепу и первым шейным позвонкам. Изображенный на рисунке зародыш по мере развития будет удлиняться в направлении низа страницы
Ясно, что все клетки несегментированной мезодермы не могут «включить» адгезию одновременно. Если бы это произошло, образовался бы один большой «мегасегмент» вместо необходимой серии небольших отдельных сомитов. Вместо этого эмбрион должен использовать какой-то особый механизм, чтобы сообщать коротким блокам клеток, что настал момент для изменения адгезионных свойств. При этом каждый сомит должен закончить формироваться еще до начала формирования следующего. Для выполнения этих условий клетки должны очень точно определять свое положение: находятся ли они именно в том ли месте, где должен образоваться сомит.
Уже на ранних этапах развития, когда эмбрион был еще очень мал, клетки по-разному реагировали на сигналы в зависимости от места и времени, например при создании первичной полоски. Например, во время гаструляции эмбрион был достаточно мал, чтобы градиент концентрации сигналов от ПВЭ определял примерную ориентацию оси «голова – хвост». Однако по мере роста эмбриона простой механизм градиентов концентрации стал все меньше подходить для разметки сложных структур организма. Чем длиннее ткань, которая должна быть размечена с помощью одного концентрационного градиента, тем более пологим будет градиент относительно размера любой клетки (рис. 23). Чем градиент более пологий, тем меньше разница концентраций для соседних клеток. Соответственно, клетке труднее «принимать решения», связанные с ее точным местоположением в эмбрионе. Системы концентрационных градиентов было вполне достаточно, чтобы наметить «черновые» различия поведения клеток раннего эмбриона, однако она просто не справится с задачей нанесения точной детальной разметки на тело, имеющее гораздо большую площадь.
Эмбрион решает эту проблему, не отбрасывая идею градиентов окончательно: он использует короткий градиент, чтобы определить границы одного сомита. Затем этот градиент, по длине равный длине сомита, передвигается вдоль эмбриона и используется снова и снова. Механизм, с помощью которого это достигается, кажется фантастическим по своей сложности, но его существование было подтверждено многочисленными кропотливыми экспериментами над эмбрионами различных видов животных. Несмотря на то что нам еще многое предстоит узнать об этом механизме, на данный момент мы знаем достаточно, чтобы сказать, что он вполне реален.
В течение всего этого процесса клетки мезодермы, еще не сегментированной и, следовательно, находящейся позади уже сформированных сомитов, синтезируют сигнальный белок под названием FGF.[87] Они делают это по умолчанию, и каждая из клеток будет продолжать синтезировать FGF, пока что-то ее не остановит. Клетки могут обнаружить секретированный ими же FGF, и это предотвращает такие изменения их адгезионных систем, которые необходимы для формирования сомита. Другими словами, еще не сегментированная мезодерма продолжает говорить себе «еще не пора» на языке FGF.[88]

Рис. 23. Градиенты концентрации вещества, распространяющегося от источника, хорошо определяют положение клеток на небольших расстояниях, но с увеличением расстояния различия концентрации этого вещества для соседних клеток становятся незначительными. Учитывая «шум», свойственный биологическим системам, клеткам становится все труднее определить, находятся ли они внутри или же снаружи той концентрационной области, в которой их поведение должно измениться определенным образом. Фиксированные градиенты, следовательно, не подходят для разметки больших и сложных структур, формирование которых требует использования многих вариантов клеточного поведения, и не могут быть использованы для того, чтобы наметить границы между будущими сомитами эмбриона
Клетки, которые уже находятся в процессе создания сомитов, не выделяют FGF. Вместо этого они синтезируют еще одну сигнальную молекулу, ретиноевую кислоту, которая также образуется клетками в голове эмбриона. Ретиноевая кислота распространяется от последнего сомита к еще не сегментированной мезодерме. Она не может распространяться очень далеко, потому что мезодермой синтезируется молекула, которая разрушает ее. Тем не менее она может проникать в мезодерму в значительной концентрации на десятую долю миллиметра.[89] Ретиноевая кислота останавливает синтез FGF в мезодерме. Таким образом, образуется узкое «окно», или «разрешающая зона», в еще не сегментированной мезодерме, в котором не хватает FGF, чтобы предотвратить образование сомита (рис. 24).

Рис. 24. Противоположно направленные градиенты ретиноевой кислоты (РК) и FGF. Градиент РК имеет максимум на головном конце тела, а FGF – на хвостовом конце. Эти градиенты задают узкую «разрешающую зону», в которой клетки могут создавать новый сомит
Определение границ этого окна в пространстве является очень важной частью механизма, в ходе которого новые сомиты образуются один за другим, но этого пока недостаточно, чтобы решить проблему. В конце концов, если клетки смогут превратиться в сомиты, как только разрешающая зона перемещается достаточно далеко к хвосту, они будут делать это одна за другой. Никогда у каждого из сомитов не будет четкой задней части, потому что всегда другие клетки, избавившись от влияния ингибиторов, попытаются сказать «и я тоже!». Такая система не сможет разделить ткани на блоки, и все, что получится, это очень длинный, бесформенный «мегасомит». Однако эту проблему можно решить, если клетки не будут формировать сомит, пока разрешающая зона не переместится на расстояние, равное длине сомита, к хвосту.[90] Тогда все клетки в этой зоне будут иметь возможность участвовать в процессе. Но это возможно только тогда, когда разрешающая зона переходит на расстояние, равное длине следующего сомита, и так далее. Кажется, именно так устроена эта система в действительности: переключение между клетками в разрешающей зоне регулируется «тиканьем» молекулярных часов.
Эти «часы» состоят из сети белков, взаимно контролирующих синтез друг друга. В некоторых случаях они, прямо или косвенно, подавляют собственный синтез. Чтобы понять, как контролируемый синтез белков может превратиться в часы, стоит рассмотреть очень простую «игрушечную» систему. Она намного проще, чем в случае реальных сомитов: эта система состоит из одного-единственного гена, в последовательности которого закодирован белок, который синтезируется в момент активности гена и его же дезактивирует. Другими словами, белок подавляет собственный синтез. Будем считать, что подобный ген присутствует в клетках, так что работа основных биохимических процессов, связанных с синтезом белка, нам уже гарантирована. Напомним, что с активных генов сначала транскрибируются РНК – «копии». Затем с этой РНК в процессе трансляции синтезирует необходимый белок (глава 1).
Предположим, что процессы транскрипции и трансляции происходят за очень короткое время по сравнению со временем жизни РНК и молекул белка. Тогда в нашей системе-«игрушке» концентрация белка постепенно выйдет на стабильный уровень. Если в начале работы системы концентрация белка ниже этого уровня, активность гена не будет сильно подавляться. РНК будет синтезироваться и транслироваться, а концентрация белка при этом вырастет. Появится больше молекул белка, которые будут ингибировать активность гена, и производство новых белковых молекул будет снижаться до тех пор, пока не выйдет на уровень, позволяющий лишь компенсировать потерю белка, связанную с окончанием срока жизни молекул (рис. 25, а). Если же концентрация этого белка изначально слишком высока, работа гена будет подавляться, и новые белки не будут синтезироваться до тех пор, пока исходная концентрация белка не понизится за счет старения и распада молекул (рис. 25, б). Основная функция этой сетевой системы состоит в том, чтобы поддерживать концентрацию белка в четко заданных пределах. Для белков-долгожителей (срок их жизни измеряется в часах и днях) небольшое запаздывание в работе системы, связанное с паузами на транскрипцию и трансляцию (на них уходят минуты), не имеет большого значения. Такие системы используются клетками для поддержания количества отдельных молекул в пределах допустимого.

Рис. 25. Поведение системы, в которой белок (обозначен жирной линией) подавляет синтез собственной РНК (обозначена тонкой линией). В результате возникает запаздывание, незначительное по сравнению со временем жизни белка. Даже если исходная концентрация белка слишком низкая (а) или слишком высокая (б), она все равно придет к стабильному значению. Сравните этот рисунок с рис. 26
Если теперь предположить, что белок «живет» так недолго, что время, необходимое для его создания, сопоставимо со временем его жизни, поведение системы становится совершенно другим. Рассмотрим ситуацию, когда в нашей модельной системе концентрация белка очень низкая. В присутствии небольшого количества белка транскрипция гена не подавляется и может проходить быстро. С некоторым запаздыванием увеличивается концентрация РНК, а немного позже, по мере трансляции РНК, вырастет и концентрация белка. С увеличением числа молекул белка транскрипция гена подавляется, но, пока концентрация РНК достаточно высока, белок будет производиться за счет трансляции этой РНК до тех пор, пока она не деградирует. В результате концентрация белка превысит «пороговый» уровень (рис. 26). По-настоящему высокой концентрации белка достаточно, чтобы полностью выключить транскрипцию гена. По мере того как молекулы РНК и белка деградируют, концентрация белка уменьшится, и транскрипция начнется снова. Однако из-за запаздывания, связанного с транскрипцией и трансляцией, понадобится некоторое время, прежде чем концентрация белка начнет расти. Быстрое разрушение белка говорит о том, что его концентрация резко уменьшается и «пороговый» уровень опять будет перейден, но уже в обратном направлении. В системе опять становится слишком мало белка, а это значит, что цикл начнется снова. Запаздывания, связанные с транскрипцией и трансляцией, вызывают многократные резкие скачки системы: от низких значений концентрации к высоким. В результате вместо стабильности мы получаем чередование высоких и низких уровней концентрации белка. Это напоминает тиканье часов.
Сетевая система «часов сегментации» значительно сложнее, чем один белок, контролирующий собственный синтез.[91] Судя по всему, функциональное ядро этих часов основано на системе реакционных петель с отрицательной обратной связью (это означает, что их компоненты репрессируют сами себя), похожих на те, что уже были описаны. Они зависят от белков, время жизни которых коротко по сравнению со временем, которое затрачивается на транскрипцию и трансляцию, необходимые для их синтеза. В работу часов вовлечено большое количество белков. Они способствуют стабильной работе часов и «сверяют» часы соседних клеток. У мышей полный цикл «часов сегментации» занимает около двух часов. Одна фаза этого цикла дает разрешение любой клетке, находящейся в разрешающей зоне еще не сегментированной мезодермы (с низким содержанием FGF), принять участие в создании сомита. Идет чередование фаз. В короткую фазу часов («тик») клетки в окне с малым содержанием FGF могут стать клетками сомита. В долгую фазу («так») они не участвуют в образовании сомита, даже если находятся в пределах разрешающей зоны. Такое чередование означает, что у разрешающей зоны есть запас времени для продвижения к хвостовой части на длину сомита до того, как клетки, обнаружившие, что они находятся в этой зоне, смогут поучаствовать в создании сомита.

Рис. 26. Поведение системы, в которой белок подавляет синтез собственной РНК. Запаздывание в системе достаточно велико относительно времени жизни белка. В начале рассматриваемого периода исходно высокая концентрация белка понижается. При этом концентрация РНК низкая. Концентрация короткоживущих белков продолжает падать, и хотя на данный момент она и так слишком мала, чтобы ингибировать синтез РНК, понадобится довольно много времени для накопления РНК. Концентрация белка опускается ниже среднего уровня, и благодаря этому РНК синтезируется в больших количествах и аккумулируется. За счет повышения концентрации РНК концентрация белка тоже постепенно увеличивается и переходит средний уровень еще до того, как белку удается подавить синтез новой РНК. В этой модели скачки многократно повторяются. Так и создается некое подобие осциллятора, или «часов»
Затем мезенхима образует блоки, длина которых определяется протяженностью разрешающей зоны и частотой тиканья часов (рис. 27). Рассмотренный здесь механизм работы часов приводит к ряду интересных эффектов: если бы часы шли быстрее, разрешающая зона не продвинулась бы так далеко в хвостовую часть. Это привело бы к возникновению большего количества более мелких сомитов. У таких животных, как змеи, гораздо больше позвонков, чем у людей. Эмбрионы змей создают больше сомитов. Недавно ученые исследовали часы сегментации в эмбрионе змеи. Оказалось, что они работают гораздо быстрее, чем у мышей или людей. Все соответствует вышеизложенной теории.[92]
Четыре механизма действуют на локальном, клеточном, уровне. Производство ретиноевой кислоты, производство FGF, часы сегментации, рост эмбриона по направлению к хвосту – все это позволяет интегрировать пространство и время так, чтобы клетка могла «решить», участвовать ли ей в создании нового сомита. У клеток нет карты внутреннего строения эмбриона. Они даже не знают собственного места нахождения. Мы опять сталкиваемся с ситуацией, в которой клетки могут самоорганизоваться в сложно устроенные структуры достаточно большого пространственного масштаба, используя только набор локальных правил.
Описанная выше система решает проблему деления мезенхимы на серию сегментов. Это не решает проблему создания на основе сомитов структур, которые не одинаковы на разных уровнях передне-задней оси, скажем позвонков в организме человека. Сомиты в грудной области, например, специализируются на формировании позвонков среднего размера, к которым крепятся ребра. Сегменты поясничного отдела формируют крупные позвонки. К ним ребра уже не крепятся! Сомиты в области крестца формируют мелкие позвонки, которые сливаются воедино, чтобы образовать заднюю часть костяного кольца, которая защищает область таза. В связи с этим клетки каждого сомита должны «знать», в какой области организма они находятся. После многолетних генетических исследований ученые начинают догадываться, каким образом клетки это «понимают». По нашим нынешним соображениям, эта система использует время, переводя пространственную информацию с молекулярного уровня на уровень эмбриона как целого. Безусловно, по мере дальнейших исследований наши представления о работе этой системы могут и измениться.

Рис. 27. Принятая в настоящее время модель использования времени и пространства для разделения непрерывного слоя ткани на сомиты. Диаграмма слева, как на рис. 24, отображает заднюю часть эмбриона сразу после формирования новой пары сомитов. Ретиноевая кислота (РК), синтезированная более ранними сомитами, распространяется к хвосту и постепенно расходуется. На диаграмме это показано заштрихованными перевернутыми треугольниками. Белок FGF, синтезируемый в хвостовой почке, распространяется к голове и также постепенно уходит (на диаграмме это отмечено треугольниками). Соотношение РК к FGF варьируется на разных участках оси «голова – хвост» мезенхимы вплоть до последних сомитов. Если клетки находятся в определенном диапазоне соотношения РК к FGF, они могут участвовать в формировании сомита. «Разрешающая зона» отмечена на схеме черной звездочкой. С течением времени рост хвостовой части эмбриона продолжается. Созревающие сомиты начинают синтезировать РК. Так «окно», определяющее клетки, которые будут создавать новый сомит, постепенно продвигается к «хвосту». Клетки в этой зоне возьмутся за формирование сомита только тогда, когда часы находятся в фазе «тик». К тому времени, когда случится следующий «тик», клетки, создававшие сомит, завершат свою работу. Так вырисовывается четкая граница между старыми и новыми сомитами. Затем процесс повторяется столько раз, сколько нужно, чтобы создать все сомиты тела
Необходимая пространственная информация хранится на молекулярном уровне в самой ДНК в виде четырех генных кластеров. Типов так называемых HOX-генов всего тринадцать. Их так и принято нумеровать: от 1 до 13. В каждом из четырех кластеров – HOXA, HOXB, HOXC и HOXD – HOX-гены, расположены в порядке нумерации (рис. 28). Таким образом, ген типа 1 в кластере HOXA называется HOXA1, ген типа 1 в кластере HOXB – HOXB1. Следовательно, в кластере HOXA гены располагаются в порядке HOXA1, A2, A3 и т. д. Ни в одном кластере нет версии каждого гена от 1 до 13. Таким образом, если положение генов изобразить на диаграмме, поместив гены одного типа в одну колонку, в каждом HOX-кластере будут пробелы (рис. 28). Есть весомые свидетельства того, что у наших далеких предков был только один HOX-кластер (именно этот кластер по-прежнему есть у насекомых, например у мучного хрущака), но в процессе эволюции позвоночных HOX-кластер был скопирован дважды. Так образовались первые два кластера (как у бесчелюстные позвоночных), а затем и еще два (как у челюстноротых позвоночных).[93] Это объясняет, почему HOXA1 так похож на HOXB1, а HOXA2 так напоминает HOXB2 и т. д. С тех пор каждый кластер утратил некоторые гены. Предположительно это случилось потому, что вскоре после дублирования гены были настолько похожи, что любой из них мог заменять другой. Потеря предками гипотетического H OX D 6, например, не имела значения, если H OX A 6, H OX B 6 и HOXC6 могли выполнять все его функции. Со временем однотипные гены разных кластеров мутировали и приобретали значительные функциональные отличия, а значит, дальнейшие потери стали невозможны. Верить или не верить таким историям об эволюции – решать вам. Факт в том, что человеческие HOX-кластеры теперь имеют такой вид (рис. 28).
Поразительно, что в течение каких-то 460 млн лет челюстноротые позвоночные плавали, ходили или летали, а порядок HOX-генов внутри кластера ни разу не менялся! При этом большинство других генов уже пережили огромное количество перемен и многократно меняли взаимное расположение. На сегодняшний день последовательность генов у разных типов животных отличается. Причины такого постоянства расположения HOX-генов в кластере напрямую связаны с порядком специализации сегментов вдоль оси тела.

Рис. 28. Генетическая структура четырех человеческих HOX-кластеров. Каждая горизонтальная линия обозначает непрерывную область хромосомы. Каждая ячейка с номером – ген HOX (например, ячейка 1 кластера HOXA – HOXA1 и т. д.)
Генные HOX-кластеры начинают проявлять активность в структурировании организма уже во время гаструляции. На каждом участке оси от шейного отдела к поясничному эта активность проявляется незадолго до образования сомитов. В то время как клетки в ходе гаструляции продвигаются через узелок, активируются гены с левого конца HOX-кластеров («левый» на рис. 28). Процесс начинается с будущего шейного к поясничному отделу. Позже, когда клетки, направленные на формирование задней части тела, вовлекаются в гаструляцию, активируются и гены, расположенные правее в HOX-кластерах. Еще через некоторое время, когда гаструляция продвинется дальше по оси «голова – хвост», активизируются гены, изображенные на рис. 28 правее. В каждом случае клетки «запоминают» набор HOX-генов, которые они «подключили», как только появились из узелка. Они в течение долгого промежутка времени поддерживают активность этих генов. Возможно, не стоило так упрощать, но в данном случае это даже полезно.
Таким образом, клетки, которые проходят стадию гаструляции раньше, «включают» HOX-гены слева (см. рис. 28). Клетки, подвергшиеся гаструляции позже, активируют HOX-гены справа. Как это можно контролировать? Есть способы. Можно направить процесс активации потенциальных генов направо через кластеры HOX-генов. Клетки в это время ожидают гаструляции. Согласно этой идее, когда клетка ныряет через узелок, чтобы гаструляция произошла, она на самом деле активирует экспрессию множества HOX-генов, которые потенциально могут быть задействованы в нужный момент. Она помнит этот набор. Клетки, которые гаструлируют позже, находятся дальше от будущей головы эмбриона, экспрессируют набор генов дальше вправо (см. рис. 28). Так волна активации потенциальных генов будет иметь больше времени для продвижения направо через кластер. По этому механизму, порядок генов в HOX-кластерах на молекулярном уровне транслируется на порядок экспрессии генов HOX на эмбриональном уровне. А эмбрион уже достиг миллиметрового диапазона. Это один из немногих примеров взаимосвязи генетической и эмбриональной структуры.
Прежде чем изучать последствия экспрессии генов на разных уровнях оси «голова – хвост», стоит потратить немного времени, чтобы выяснить, как волна активации потенциальных генов в первую очередь проходит через HOX-кластеры. Детали этого процесса до сих пор тщательно исследуются, но общая схема механизма, кажется, начинает вырисовываться сквозь дымку множества экспериментальных результатов. В хромосомах ДНК может быть довольно компактно упакована среди белков. Это полезно для экономии пространства, но при этом гены недоступны для транскрипции. Принципиально ДНК может быть и в свободной, легко доступной форме. Большую часть времени многие участки хромосом плотно упакованы, а более свободные «петли» ДНК выходят за их пределы. Некоторые участки упакованы плотно. Некоторые никогда не были так упакованы. Некоторые из них могут быть в любом состоянии, в зависимости от присутствия того или иного ДНК-связывающего белка. Последние данные, полученные в ходе экспериментов над мышиными эмбрионами, позволяют предположить, что по крайней мере один из hoх-кластеров,[94] hoxb, может быть в любом из обозначенных состояний.
Перед активацией hoxb-генов кластер в компактном состоянии. Из всех генов hoxb-кластера самый правый ген (hoxb13) «похоронен» в глубине компактного клубка. hoxb1 наиболее доступен, будучи в начале компактной зоны. Экспрессия генов hoxb зависит от освобождения из этой «компактной тюрьмы». Это может случиться, если специфичные последовательности ДНК-связывающих белков присоединятся к ДНК с наиболее доступного конца и «перепрограммируют» упаковочные белки, так чтобы они позволили ДНК принять более свободную, петлеобразную форму. Эти ДНК-связывающие белки активируются ретиноевой кислотой. Эффект ретиноевой кислоты на hoxb-кластер настолько мощный, что его можно разглядеть даже в мышиных СКЭ, выращенных в простой чашке Петри.[95] Если постепенно добавить туда небольшое количество ретиноевой кислоты, «спрессованный» hoxb-кластер начинает ослабевать с конца. Так что hoxb1, hoxb2, hoxb3 и так далее будут доступны для транскрипции (рис. 29). Все это требует времени, что не удивительно, учитывая, что в ДНК hoxb-кластера около 150 000 оснований. Освобождение HOXb гена – постепенный процесс. Необходимо осуществлять контроль над порядком их «укладки» в кластере. Вероятно, то же самое верно[96] для hoxa-, с – и d-кластеров мыши и, по аналогии, для всех кластеров HOX человека.
Получение петли из hoxb-кластера также наблюдается в эмбрионе мыши. Скорость «разматывания» и волны активации генов совпадают с той скоростью, с которой гаструляция распространяется по направлению к «хвосту».[97] Таким образом, можно смело надеяться на то, что описанный выше механизм верен, хотя мы уже знаем, что его сопровождает огромное количество дополнительных сложностей. Необходимо убедиться, что клетки «запоминают» те гены для экспрессии, и это должно быть согласовано с соседними клетками. При этом отключаются гены слева, а те, что справа, подвергаются экспрессии. Тщательная систематизация всего этого процесса может стать делом жизни многих исследователей.
Человеческие HOX-гены (как и мышиные hox-гены) очень важны, поскольку существует связь между набором HOX-генов, экспрессированных сомитом, и типом будущего позвонка.[98] Например, третий, четвертый и пятый позвонки шейного отдела очень похожи: образовавшие их клетки экспрессируют тот же набор генов. Следующие два позвонка от них отличаются, и их клетки уже экспрессируют другие наборы HOX-генов.

Рис. 29. Постепенное изменение структуры HOX-кластера, от компактной до более рыхлой, высвобождает большее количество генов для транскрипции в том порядке, в котором они расположены в кластере
У различных видов животных число позвонков варьируется. У кур, например, четырнадцать шейных позвонков, а у мышей, как и у нас, – семь. В обоих случаях тем не менее начало экспрессии генов типа HOX6 (Hoxa6, Hoxb6 и т. д.) знаменует конец сомитов, создающих шейные позвонки, и начало тех, которые образуют позвонки грудного отдела. Поэтому гены HOX коррелирует с типом позвонков, а не просто с их числом. Аналогично начало экспрессии генов типа HOX10 означает конец сомитов, идущих на грудные позвонки, и начало тех, которые создают позвонки поясничного отдела. Гены типа HOX11 отвечают за переход от поясничного к крестцовому позвонку. Все это подтверждает, что тип HOX-генов может определять вид позвонков, который создадут сомиты.
Эта идея была проверена экспериментально в ходе генной инженерии эмбрионов мыши. Из них убирали один или несколько Hox-генов и наблюдали последствия этой операции для развития. В шейном отделе нормальной мыши должен быть один первый позвонок («атлант») и один второй («ось»). У каждого из них своя специфика, они позволяют по-разному осуществлять кивки и вращения головы. У третьего, четвертого и пятого позвонков форма, характерная для позвонков шейного отдела. При удалении гена Hoxa4 третий, неспециализированный, позвонок развивается как второй, осевой. Клетки как будто «думают», что они ближе к головной части зародыша. Кроме того, у мышей без генов Hoxa7 и Hoxb7, отвечающих за первые позвонки грудного отдела, позвонки, к которым должны крепиться ребра, развиваются как позвонки шеи, без ребер. Опять же, без этих генов клетки ведут себя так, как если бы они были ближе к голове, чем на самом деле. С другой стороны, когда удаляются Hoxa5 и Hoxa6, клетки в той же области тела «думают», что должны располагаться дальше от головы, где эти гены, естественно, дезактивируются. В результате седьмой позвонок шейного отдела имеет ребра, как полагается первому позвонку грудного отдела. То же происходит и ниже, если гены по правую сторону (см. рис. 28) удаляются.
В обеих системах сегментации, будь то «часы» и градиенты, благодаря которым тело делится на сомиты, или система HOX-генов, сообщающая клеткам этих сомитов специализацию, в масштабах организма протекают простые локальные процессы. В обоих случаях в этих процессах используются определенные временные промежутки для создания структур в пространстве. Это еще один пример того, как сложные клеточные системы эмбрионального развития пользуются простейшими математическими принципами и создают нечто гораздо большее, чем они сами.
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги