2. Эпизоотология и эпидемиология чумы
2.1. Некоторые аспекты природной очаговости чумы
Чума является типичной зоонозной инфекцией. К настоящему времени она выявлена более чем у 200 видов грызунов и заячьих [Kirsch A., 1986], а также у других животных, включая слонов и буйволов, медведей, верблюдов, коз, собак и кошек. Перечень видов животных, которые втягиваются в эпизоотии, можно найти в работах Wu Lien-Teh и соавт. [1936], R. Pollitzer [1954], Ю. М. Ралля [1958], В. М. Туманского [1958]. Птицы, пресмыкающиеся и земноводные к чуме не восприимчивы [Федоров В. Н. и др.,1955].
Цепь, состоящую из возбудителя (Y. pestis), носителей (грызунов) и переносчиков (блох), принято называть «триадой», а определенные географические ландшафты, заселенные грызунами, видовые и межвидовые отношения которых обеспечивают непрерывную циркуляцию чумного микроба от донора через членистоногих реципиентам, — природными очагами чумы.
Е. Н. Павловский [1946] подчеркивал, что все соотношения в биогеоценозе между возбудителями природно-очаговых заболеваний, их донорами, переносчиками и реципиентами сложились в процессе эволюции организмов и межвидовых отношений на определенном фоне внешней среды без какой-либо зависимости или связи его с человеком, а может быть, для некоторых болезней еще ранее появления человека на Земле. Однако влияние человека может перестраивать «патобиоценозы», а иногда даже укреплять и расширять их («антропургические» очаги).
Признание природного очага «…пространственно ограниченным, исторически сложившимся естественным комплексом организмов создало базу для научного изучения двух основных сторон этого явления: 1) биологической структуры очага или механизма и условий взаимодействия его компонентов; 2) пространственной структуры очага и взаимодействия его отдельных частей, заселенных разными популяциями возбудителя и его хозяев…» [Наумов Н. П. и др., 1972].
С позиций учения Е. Н. Павловского между «природными очагами» и «очагами инфекций» имеются принципиальные отличия. Первые являются постоянными, существующими «вечно», вторые — эфемерны.
Укоренение мнения о неравноценности в эпизоотологическом отношении территории любого очага привело к появлению представлений об автономных очагах как части «категории более высокого ранга» — природного очага и микроочагах — наименьшей структурной единице очага, лежащей «в основе эпизоотологии чумы, механизмы которой сейчас неизвестны» [Дятлов А. И., 1989].
В русскоязычной литературе принято делить всех носителей инфекции на основных и второстепенных. Последних, в свою очередь, подразделяют на случайных и факультативных [Фенюк Б. К., 1948]. Наряду с этим предлагается различать: 1) основные резервуары инфекции в очаге; постоянные (основные хозяева) — главное «место» пребывания возбудителя и временные, в популяциях которых возбудитель размножается периодически — в годы подъема численности или в определенные сезоны; 2) распространителей инфекции и её переносчиков — высокоподвижные виды или виды, склонные к массовому размножению и широким миграциям [Наумов Н. П. и др., 1972]. По нашему мнению, предложение Н. П. Наумова и соавт. по духу больше соответствует тем представлениям об эпизоотологии чумы, которых придерживаются зарубежные учёные [Pollitzer R, 1954; Butler T., 1983].
Согласно устоявшимся взглядам, особая роль основных носителей в природных очагах чумы обусловлена наличием у них общих экологических признаков: относительно высокой и, главное, относительно устойчивой численности сочленов популяции и паразитированием на них таких видов блох, которые могут быть активными переносчиками чумы и обладают большой жизнеспособностью в условиях нор и гнезд грызунов. Второстепенные носители, как правило, этими признаками не обладают [Федоров В. Н. и др., 1955; РалльЮ. М., 1965].
Помимо перечисленных особенностей основных носителей большое значение придается обязательному наличию у них на тех или иных стадиях инфекционного процесса бактериемии — основного условия трансмиссивного механизма передачи инфекции [Федоров В. Н.и др., 1955; Ралль Ю. М., 1965; Наумов Н. П. и др., 1972; Козлов М. П., 1979;Pollitzer R, 1954; Baltazard M. et. al, 1953], хотя эта особенность должна быть присуща также второстепенным носителям. Однако необходимо отметить, что четкую границу между основными и второстепенными носителями чумы провести не всегда возможно. Причины этого будут ясны из дальнейшего изложения.
Логическим следствием деления носителей на основных и второстепенных (или «основных», «временных» и «распространителей») стал постулат о моногостальности очагов — «второй закон природной очаговости чумы» [Ралль Ю. М., 1965]. Согласно этому положению в каждом природном очаге есть свой основной носитель чумной инфекции, тогда как другие грызуны, хотя и вовлекаются в эпизоотии, но в поддержании очага роли не играют. При этом допускается, что носитель, являющийся в одном очаге «основным», в другом может переходить в разряд «второстепенных» [Ралль Ю. М., 1965].
Как весомый аргумент в пользу моногостальности очагов исследователи приводили факт связи между видом основного носителя и характерными свойствами возбудителя эпизоотии в его популяциях, что послуживо основанием для создания различных внутривидовых классификаций Y. pestis (см. раздел 3.4.). Одним из первых такую связь подметил В. М. Туманский [1958], предложивший различать три разновидности Y. pestis: «крысиную» (ratti), «сурчиную» (marmotae) и суслиную» (citelli). Сторонники этой точки зрения утверждали, что особенности штаммов чумного микроба из разных природных очагов являются отражением взаимной адаптации микроба и его основных хозяев, следствием их коэволюции [Ралль Ю.М., 1965; Козлов М. П., 1979; Дятлов А. И., 1989]. По их мнению, «взаимная адаптация» привела к изменению ферментативной активности или потребности в источниках питания у возбудителя чумы и появлению у него избирательной вирулентности. Адаптация же носителей сказалась на их чувствительности к чумной инфекции. Предполагается, в частности, что «…малая чувствительность вида указывает на связь и постоянный контакт этого вида и его ближайших предков с возбудителем чумы. Наоборот, высокая чувствительность говорит о том, что в процессе эволюции этот вид недавно встретился, совсем не встречался или имел слабый контакт с возбудителем чумы» [Кучерук В. В., 1965].
Наряду с ярыми приверженцами теории моногостальности имеется немало ученых, которые отвергают ее или разделяют частично.
Как считали Н. И. Калабухов [1949], Н. В. Некипелов [1959а], В. А. Саржинский [1969] и некоторые другие, очаги чумы — полигостальны. А. А. Лавровский и С. Н. Варшавский [1970] полагали, что взгляды на структуру природных очагов как на моногостальные системы нельзя полностью заменить полигостальной концепцией. Они допускали одновременное существование поли- и моногостальных очагов как в эволюционно-историческом аспекте, так и в современных условиях, хотя критериев деления очагов чумы на моно- и полигостальные не приводили. В. П. Хрусцелевский [1974] предлагал вообще отказаться от определения гостальности очагов. По его мнению, хозяевами чумного микроба являются не только грызуны, но и блохи, поэтому все очаги в принципе полигостальны.
Представление о природной очаговости инфекций вообще и чумы в частности разделяется за рубежом [Pollitzer R, 1954; Объединенный комитет экспертов ФАО/ВОЗ по зоонозам, 1969; Комитет экспертов ВОЗ по чуме, 1971]. Вместе с тем, там чаще используется термин «очаги дикой чумы» (wild plague), противопоставляемый термину «очаги домовой чумы» (domestic plague), которые связаны с наличием синантропных грызунов, главным образом с крысами (Rattus rattus и R. norvegicus). Последние также называют «крысиными», «городскими» или «портовыми». В этой связи уместно сделать одно замечание. Оно касается утверждения Ю. М. Ралля [1965] о том, что «собственно природных очагов чумы на крысах не бывает и под «зоной крысиной чумы» следует понимать область наиболее частых и упорных вторичных эпизоотий среди крыс». Однако подобное утверждение идёт вразрез с многочисленными фактами, особенно ставшими известными относительно недавно [Козлов, 1979; Вильямс Д. И. и др., 1980; Козакевич В. П. и др., 1981; Акиев А. К. и др., 1983;, 1984; Pollitzer R., 1954; Butler T, 1983; Tomich P. F. et. al.]. В то же время существуют и «крысиные» очаги в понимании Ю. М. Ралля, хотя, как подчеркивали еще В. Н. Федоров и соавт. [1955] «эти связи не всегда удается точно установить».
Природные очаги чумы разбросаны по всему свету и располагаются между 48–49° северной и 40–41° южной широты. Несмотря на это, сейчас чума редко передается людям и её вспышки чаще носят ограниченный характер. Представление об «эпидемическом потенциале» природных очагов на исходе XX в. можно получить из данных, приведенным в табл. 1 и 2. Однако в этом контексте, пожалуй, наиболее полезны сведения об очагах чумы в СНГ, где они занимают свыше 2 млн. км2 и эпизоотии регистрируются до сих пор, хотя случаев заболевания людей за последние 60 лет было относительно не много (табл. 3). Впрочем, не следует думать, что «чума перестала быть чумой». При ослаблении внимания к очагам они в любой момент могут выйти из-под контроля, что может вызывать серьезные эпидемические осложнения, как это было в Маньчжурии (1910–1911, 1930–1931 и 1946–1947 гг.), Бирме (1974 г.) или во Вьетнаме (в 60-70-х годах).



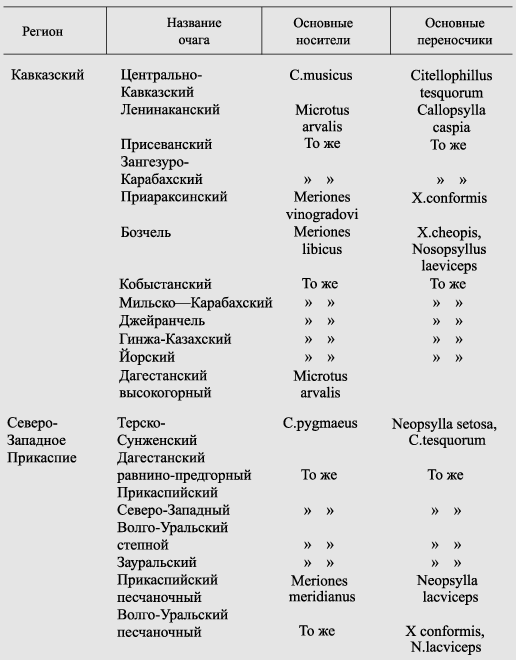
Оставив в стороне споры о гостальности, приведем краткую характеристику природных очагов в СНГ (табл. 4). Сведения об эпизоотологии чумы среди основных носителей чумы в этих очагах приводятся в одной из следующих глав. Здесь же мы остановимся на одном из очагов Юго-Восточной Азии — «колыбели» третьей пандемии чумы, а именно на вьетнамском. Это связано с тем, что в течение почти 10 лет, начиная с 1965 г., на его долю приходился пик всех заболеваний чумой.

Примечание. Общая площадь очагов около 2,1 млн км2.
Как указывали А. К. Акиев с соавт. [1983], характерной особенностью эпизоотий чумы во Вьетнаме следует считать то, что инфекция стойко удерживается в определенных районах: на равнинах северо-западнее и западнее Сайгона в провинциях Тайнинь и Хаунгхиа; на Центральной приморской равнине (провинция Тхуанхай); на Центральном плоскогорье (плато Зилинь, Ма Дарлак); в северной приморской области бывшего Южного Вьетнама от прибрежных районов до подошвы горных отрогов Аннамского хребта, на которые приходилось почти 90 % всех случаев чумы.
А. К. Акиев и соавт. считали, что природные очаги чумы на плоскогорьях Вьетнама смыкаются с очагами в Кампучии и Лаосе.
Во Вьетнаме установлена несомненная роль в эпизоотиях чумы трех видов крыс (чёрной, серой и малой (R. exulans), а также крупной землеройки Suncus murinus [Marshall J. D. et al., 1967]. Из числа грызунов эпидемиологическое значение серой крысы особенно велико в приморских селениях, а черные крысы имеют основное значение на плато. Помимо указанных видов животных, чума неоднократно выявлялась у других видов крыс, бандикотов, в частности, B. indica, и домовых мышей.
Основным, если не единственным, видом переносчиков во Вьетнаме является блоха X. cheopis (на её долю приходится 99 % всех блох, снимаемых с грызунов).
Близкая к Вьетнаму ситуация по чуме отмечается на Яве [Вильямс Д. И. и др., 1980] и в Бирме [Козакевич В. П. и др., 1984].
С эпидемиологической точки зрения наиболее опасны стыки между очагами дикой чумы и поселениями людей, где возбудитель передается синантропным грызунам — крысам и домовым мышам, а также домашним животным.
В. П. Козакевич и соавт. [1981], не исключая «возможности и дополнительного укоренения чумы в популяциях городских крыс вследствие заносов её в портовые города», ставят вопрос о том, насколько реально при этом возникновение новых природных очагов (новой энзоотии) — Это (не праздный вопрос, поскольку с заносами чумы могло быть связано появление её природных очагов там, где их никогда не было.
Многие авторы, в частности, Д. И. Вильямс и соавт. [1980], L. G. Lipson [1972], T. Butler [1983], P. F. Tomich и соавт. [1984]. считают, что чума в США и Индонезии, на Гавайских островах и Юге Африки укоренилась после завоза её в портовые города в ходе третьей пандемии. С этих позиций примером новых природных очагов надо считать Запад США, где носителями чумы являются грызуны, принадлежащие не менее, чем к 10 родам, два вида из семейства заячьих [Meyer K, 1960] и даже дикие плотоядные [Butler T., 1983]. Однако с этим согласны не все, в том числе В. Н. Федоров и соавт. [1955], Ю. М. Ралль [1965] и K. Meyer [1960], отстаивавшие точку зрения об автохтонности всех природных очагов чумы. В какой-то мере аргументом в пользу последнего мнения, если иметь в виду США, могут служить следующие факты. В период интенсивных заносов чумы в 1900–1909 гг. в Калифорнии было 332 случая «портовой» чумы, а в последующем на Западе США регистрировались только случаи «дикой» чумы, пик которых (105 заболеваний, пришелся на период 1970–1979 гг. [Butler T., 1983]. Впрочем, приведенные данные можно трактовать и как результат повышения активности природных очагов, становление которых продолжается до сих пор. Иначе чем объяснить необычную для очагов Старого света полигостальность американских очагов — Так или иначе, но несмотря на огромные масштабы завоза в первой трети нашего века [Николаев Н. И., 1968], чума укоренилась далеко не везде. Классическим примером является Австралия — единственный континент, по-прежнему свободный от чумы. Ярый сторонник идеи об автохтонности всех природных очагов чумы, Ю. М. Ралль, полагал, что появление чумы как зооноза было связано с развитием на Земле высших плацентарных млекопитающих, а Австралия отделилась от Евро-Азиатского материка значительно раньше и потому заселена клоачными и сумчатыми животными; мышевидные грызуны — потенциальные носители чумы, там весьма малочисленны. Еще одна причина может заключаться в том, что в Австралии очень низка численность блохи X. cheopis, играющей основную роль в поддержании «крысиной» чумы. Именно «дефицит» переносчиков чумы [Cumpston, цит. по Wu Lien-Teh et al., 1936] избавил Австралию от укоренения в этой стране «заносной» чумы. Подтверждением такой точки зрения служит то, что чумы никогда не было на Тасмании, где X. cheopis вообще нет, и более интенсивные, по сравнению со штатом Виктория, вспышки завезенной чумы в Квисленде и Новом Южном Уэллсе, в которых заблошивленность крыс была значительно выше.
Обсуждая проблемы завоза чумы, В. Н. Федоров и соавт. [1955] указывали, что в умеренных широтах основными препятствиями для укоренения её среди местных крыс, т. е. для возникновения эндемичности, служат: резкое снижение численности блох X. cheopis в некоторые довольно продолжительные сезоны, вследствие чего нарушается передача Y. pestis от крысы к крысе; относительно низкая численность крыс и неравномерное их распределение по отдельным объектам в городах.
По-видимому, в связи именно с этими причинами чума «не укоренилась» в Одессе, несмотря на многократные завозы в прошлом веке и в начале нынешнего столетия.
Как бы подводя итоги дискуссии, Комитет экспертов ВОЗ по чуме [1971] писал: " Поскольку маловероятно, что завоз чумы морским путем приведет к возникновению новых природных очагов, желательно заменить карты районов временных проявлений чумы в прошлом картой территорий известного или предполагаемого укоренения инфекций в настоящее время и районов, в которые она может распространиться». К этому следует добавить, что речь должна была бы идти о любых путях завоза, будь то морской, воздушный или сухопутный. Однако, по нашему мнению, закрывать дискуссию еще рано.
Каждый природный очаг отличается от другого присущей ему спецификой. Эта специфика обусловливается как видовым составом носителей и переносчиков, так и ландшафтно-географическими особенностями очага. Поэтому наибольших успехов исследователи обычно достигают там, где наблюдения за очагом ведут в течение многих лет с учётом особенностей всего патогеоценоза. Примером такого подхода может служить работа уникальной по подбору специалистов противочумной системы Советского Союза, контролировавшей ситуацию по чуме по всей стране и задававшейся целью оздоровить её природные очаги [Наумов Н. П. и др., 1972; Голубинский Е. П. и др., 1987; Fenuk B. K, 1960; Kuceruk V. V., 1970].
Одной из важнейших черт любого природного очага является характерная для него динамика эпизоотий. Нередко они протекают вяло и в течение длительного времени. Иногда же они начинаются внезапно, «взрывоподобно», и так же быстро затухают. В таких случаях межэпизоотические периоды могут продолжаться неопределенное время, причем самые тщательные поиски часто не выявляют возбудителя чумы. Мы сами были непосредственными свидетелями пробуждения одного из очагов в районе Гулженги в Забайкалье. До этого эпизоотия там была зарегистрирована в 1947 г. Последующие многолетние исследования различных грызунов (около 300 тыс.) неизменно давали отрицательные результаты, пока, наконец, летом 1960 г. от даурского суслика, даурского хомячка, полёвки Брандта и блох не были выделены культуры, оказавшиеся, правда, слабовирулентными для лабораторных животных.
Естественно, давно уже возник вопрос о том, каким образом и где в межэпизоотические периоды сохраняется источник инфекции. К сожалению, однозначного ответа на него получить пока не удалось, о чём свидетельствует большое число гипотез [Леви М. И., 1985; Домарадский И. В. и др., 1995]. Однако некоторые авторы полагают, что межэпизоотических периодов никогда не бывает. Так, один из приверженцев этой концепции (М. П. Козлов [1970] писал: «… эпизоотический процесс пульсирует, перемещается в пространстве не только в пределах группы нор, микроочагов, участков, но и в пределах всего очага, что никаких перерывов (межэпизоотических периодов) в течение эпизоотического процесса нет,». Он выдвинул тезис об эпизоотическом процессе как о «саморегулирующейся» системе. Противоположной точки зрения придерживаются И. С. Солдаткин и Ю. В. Руденчик [1995], которые в конечном итоге вообще пришли к заключению о полной несостоятельности учения Е. Н. Павловского о природной очаговости трансмиссивных инфекций, поскольку оно зиждилось на постулате о непрерывности эпизоотического процесса, в значительной мере «подпитываемом» чумологами, сводившими всю свою работу к изучении «упрощенной системы»: грызун R блоха R грызун. Главным для И. С. Солдаткина и Ю. В. Руденчика было не наличие или отсутствие «межэпизоотических» периодов, а вопрос о возможности существования чумного микроба в природе вне грызунов или блох, поскольку, по их мнению, представление об исключительной роли «пресловутой» триады в поддержании эндемичности чумы является несостоятельным.
Комплекс проблем, относящихся к возникновению, укоренению и угасанию очагов чумы, давно уже заставили искать новые подходы к их решению. В связи с этим остановимся лишь на двух гипотезах, которые, как нам кажется, лучше других помогут в дальнейших поисках.
Более 30 дет назад, когда пробудился упоминавшийся очаг в Забайкалье, было высказано предположение о возможности существования чумного микроба в слабовирулентной форме [Краминский В. А., Домарадский И. В.]. С позиций ортодоксальных представлений о эпизоотологических процессах при чуме серьезные возражения против новой гипотезы. сводились к тому, что штаммы с пониженной вирулентностью якобы не могут циркулировать в природе [Ралль Ю. М., 1965; Козлов М. П., 1979]. Однако за последние годы накопились данные о неоднородности популяций чумного микроба по вирулентности, фенотипической флюктуации вирулентности и неоднозначной роли факторов патогенности (вирулентности) Y. pestis в эпизоотических процессах, которые позволяют не только снять указанные возражения, но и развить нашу гипотезу (см. раздел 3). Кстати, по существу, она была поддержана Комитетом экспертов ВОЗ по чуме [1971].
Вторая гипотеза принадлежит H. Mollaret [1963]. Смысл её сводится к тому, что чумной микроб при наличии соответствующих условий может длительно персистировать в почве нор («теллурическая чума»). Развивая эту гипотезу, М. Балтазар [1964] пришел к заключению, что цикл чумы в природных очагах состоит из двух фаз: паразитической (на грызунах и в их блохах — кратковременной и неустойчивой) и непаразитической (в почве нор — устойчивой и, может быть, продолжительной). В пользу концепции М. Балтазара и H. Mollaret сейчас получено так много фактов, что перечислить их здесь мы не имеем возможности. Отметим только, что найдено объяснение того, каким образом неспорообразующие микробы могут длительно существовать в окружающей среде. В данном случае речь идёт о переходе бактерий в так называемое «некультивируемое» состояние. Открытие этого принципиального нового явления избавляет от необходимости прибегать к другим, весьма сомнительным объяснениям, подобным тем, которые дает А. И. Дятлов (см. стр. 61). Не исключено, что определенную роль в сохранении чумного микроба играет также L-трансформация [Ларина В. С., 1992; цит. по Домарадскому И. В. и др.,1995].
Небезынтересно отметить, что T. Butler [1983] уже рассматривает существование чумного микроба в почве нор грызунов как непреложный факт.
Совсем недавно у автора монографии возникло предположение о возможности существования чумного микроба — типичного факультативного внутриклеточного паразита [Moulder J. W, 1985] в одноклеточных животных. О том, что оно заслуживает серьезного внимания и может служить еще одним важным аргументом в пользу гипотезы о «теллурической чуме» говорят результаты опытов C. В. Никульшина и соавт. [1992], доказавших персистенцию её возбудителя в почвенных амёбах из микробиотопов в природных очагах Центрального Кавказа, заселенных горными сусликами. Вероятность встречи этих простейших с чумным микробом, попадающим тем или иным путем во внешнюю среду, достаточно высока.
Как показали С. В. Никульшин и соавт. (1992), амебы Hartemennella rhysodes и Vahlkampfia hartmanii фагоцитируют чумной (и псевдотуберкулёзый) микроб. При этом отмечены случаи гибели вегетативных форм H. rhysodes, «наглотавшихся» клеток Y. pestis, и выход последних наружу. Выявлены также случаи сохранения чумного микроба в предцистах амеб, т. е. в условиях, в которых исключается возможность его переваривания (в предцистах нет пищеварительных вакуолей). С эпидемиологической точки зрения последний факт особенно интересен, поскольку в инцистированном состоянии одноклеточные животные выдерживают низкие температуры, обезвоживание, жару и воздействие других абиотических факторов и могут сохраняться годами. Следовательно, внутри цист чумной микроб также имеет шансы на выживание при самых неблагоприятных условиях окружающей среды и вместе с цистами может разноситься ветром на огромные расстояния. Кстати, если считать, что условия существования чумного микроба в одноклеточных организмах сходны с таковыми в фагоцитах (см. ниже), то представляется возможным дать еще одно объяснение тому, как слабовирулентные штаммы могут сохраняться в природе.
Полагаем, что все сказанное позволяет надеяться на создание новой теории очаговости чумы в природе, которая устранит теперешние противоречия в учении о ней, в частности подмеченные С. И. Солдаткиным и Ю. В. Руденчиком. Возможно, что в дальнейшем чума вообще будет рассматриваться как одна из разновидностей зоосапронозных заболеваний животных и человека.
2.2. Взаимоотношения переносчиков с возбудителем
Общепризнанно, что передача инфекции в естественных условиях от грызуна к грызуну осуществляется трансмиссивным путем, а переносчиками служат блохи. В табл. 5 представлены данные о наиболее типичных видах блох — переносчиках чумной инфекции — основных видах их прокормителей в различных зарубежных природных очагах чумы. Аналогичные сведения, относящиеся к природным очагам чумы в СНГ содержатся в табл. 4.
Блохи — мелкие бескрылые насекомые, относящиеся к отряду Siphonapthera (Aphaniptera). Этот отряд делится на два надсемейства (Pulicoides и Ceratophylloides, которые включают 17 семейств и приблизительно 1500 известных сейчас видов [Bahmanyar M., Cavanaugh D. С.,1976]. Подробности, касающиеся классификации, анатомо-физиологических особенностей и экологии блох, равно как и их роли в качестве переносчиков инфекций, можно найти в работах И. Г. Иоффа [1941], В. Е. Тифлова [1960], B. Ф. Жовтого [1966], В. А. Бибиковой и Л. Н. Классовского [1974], Wu Lien-Teh и соавт. [1936], R. Pollitzer [1954], L. Kartman и соавт. [1958], а описание их эволюции — в работах М. П. Козлова [1970] и А. И. Дятлова [1989].

Единственный способ, с помощью которого прoисходит инфицирование блох, — кровососание. Но для этого необходимо наличие бактериемии у хозяина блохи. Число блох, инфицируемых при кровососании, обычно зависит от интенсивности бактериемии. Однако при прочих равных условиях частота заражения различных видов блох неодинакова. В значительной мере она зависит от интервалов между кровососаниями блох, которые питаются редко, часто не заражаются, поскольку «пропускают» бактериемию у хозяев. У тех же видов блох, которые обычно заражаются, продолжительность сохранения микроба в их организме варьирует в широких пределах. При этом нередки случаи, когда блохи освобождаются от бактерий, что может быть связано с участием фагоцитов и действием защитных факторов насекомых, подобных «лизинам» [Штейнхауз Э., 1952] или гемагглютининам [Купер Э., 1980]. R. Pollitzer [1954] подчеркивал, что частота клиренса значительно выше у блох, являющихся низкоэффективными переносчиками инфекции, и наоборот. Известны также случаи, когда микроб в организме блох теряет вирулентность. Тогда, по мнению L. Otten [цит. по R. Pollitzer, 1954], вместо того чтобы заражать животных, блохи способствуют развитию у них иммунитета к чуме. Однако значительно чаще вирулентность чумного микроба сохраняется очень долго, иногда даже после гибели блох.
В связи с особой ролью блох как переносчиков чумы неоднократно поднимался вопрос: не болеют ли чумой они сами — Дискуссия продолжалась много лет [Козлов М. П., 1979]. Отметим кстати, что блокообразование у блох, о чём пойдёт речь ниже, К. И. Кондрашкина [1969] рассматривала как проявление чумы. Однако, если это болезнь, то она весьма своеобразна.
Вскоре после того, как в 1898 г. P. L. Simond установил роль блох в передаче чумной инфекции, стало ясно, что развитие этой способности связано с интенсивным размножением возбудителя. Его размножение бывает столь сильным, что клетки микроба, склеиваясь друг с другом, образуют комок, заполняющий весь преджелудок и даже часть пищевода блохи, вследствие чего возникает непроходимость пищеварительного тракта («чумной блок»). Несмотря на это, «блокированные» блохи не теряют потребность сосать кровь, но при наличии блока насасываемая кровь, омывающая микробный комок в пищеводе, пружинящими движениями глотки снова вгоняется в ранку на месте укуса, внося в неё большое число вирулентных бактерий. Для развития заболевания у морских свинок и белых мышей достаточно укуса одной такой блохи [Федоров В. Н. и др., 1955].
Опытным путем установлено, что иногда блок в преджелудке блохи исчезает и появляется вновь при поступлении свежей крови и благоприятной температуре и влажности. Однако не у всех блох, насосавшихся крови больных чумой животных, блок формируется в ближайшие же дни. Необходимо время, когда возникнут подходящие условия для блокообразования. Из этого следует, что чумной микроб может относительно долго сохраняться в организме блохи, причем трансовариальной передачи чумных микробов у блох нет, а личинки, как правило не заражаются (единственный известный нам случай описан Е. К. Демидовой и Н. Д. Емельяновой в 1971 г.). Небезынтересно будет узнать, что продолжительность жизни блокированных блох в общем не велика и зависит как от вида блохи, так и от климатических условий [Pollitzer R., 1954]. Например основной переносчик чумы во многих очагах X. cheopis погибает в течение нескольких дней. Но известны также случаи, когда блокированные блохи жили до 50 дней [Wu Lien-Teh et al., 1936].
В настоящее время лишь немногие сомневаются в том, что блокообразование является важнейшим фактором, ответственным за циркуляцию чумного микроба в природе, перед которым «… бледнеют все теоретические возможные механизмы передачи инфекции… [Фёдоров В. Н. и др., 1955]. Однако не всё так просто, поскольку блокообразование отмечается не у всех видов блох, выявляемых в природных очагах. Основываясь на данных литературы, R. Pollitzer [1954] объяснял это ограниченным сроком существования блока в ряде случаев или неполным соответствием опытных данных тому, что происходит в естественных условиях. Не исключено также, что у некоторых видов блох блокообразование невозможно из-за их анатомо-физиологических особенностей или не регистрируется, поскольку в природе блокированные блохи обычно быстро погибают [Дятлов А. И., 1989]. Одновременно R. Pollitzer обращал внимание на то, что даже полностью блокированные блохи не всегда заражают животных. Поэтому особого обсуждения заслуживает вопрос о способности Pulex irritans — блох человеческого жилья, быть активными переносчиками чумы. Мы акцентируем на них внимание, так как P. irritans относится к виду блох, которому присуще блокообразование. К тому же инфицированных чумным микробом P. irritans неоднократно находили в ряде активных очагов чумы земного шара, а G. Blanc и M. Baltazard [1943] удалось заразить морских свинок с помощью блох, которых кормили на больных чумой людях. Тем не менее отношение к этому виду блох как переносчикам чумы остается неоднозначным. В «крысиных» очагах чумы Китая, Индии и Мадагаскара их роль оценивается как ничтожно малая, а основное значение придается X. cheopis. В противоположность этому в «песчаночных» очагах, например в Марокко, где относительно слабая векторная способность P. irritans компенсируется относительно высокой их численностью, ей отводится главная роль. Как указывали В. Н. Фёдоров и соавт. [1955], в подобных очагах вспышки чумы обычно начинались с заражения первых заболевших непосредственно от грызунов. Затем инфекция распространялась от человека к человеку через P. irritans. Приведенные факты показывают, что помимо всего прочего один и тот же вид блох, находясь в разных условиях обитания, может изменять свою активность как переносчика чумы. К этому добавим, что сейчас лучше всего изучены блохи грызунов-комменсалов, а блохи диких животных по-прежнему остаются малоизученными. Какие сюрпризы они могут преподнести покажет будущее.
Какова же роль других кровососущих насекомых в распространении чумы? Этот вопрос обсуждался Wu Lien-Teh и соавт. [1936], R. Pollitzer [1954] и многими другими. Все они пришли к единому выводу: значение их для эпидемиологии чумы весьма невелико и «…часто кажется большим, чем оно бывает в действительности…» [Федоров В. Н. и др., 1955]. С подобным выводом нельзя не согласиться, если иметь в виду, что феномен блокообразования присущ только блохам. Несмотря на это, отдавая должное блохам как переносчикам чумы, Е. П. Голубинский и соавт. [1987] писали: «Но в учении о природной очаговости чумы есть ряд загадок, которые не решаются участием в эпизоотическом процессе одних блох. Их разгадку следует искать в исследовании всех компонентов паразитоценоза, в состав которого входят все кровососущие членистоногие — эндопаразиты носителей». Не созвучно ли с этим мнение Комитета экспертов ВОЗ по чуме [1971] о том, что хотя «…многие природные очаги чумы в умеренном климате изучены достаточно, но по тропическим районам таких данных почти нет»?
2.3. Основные сведения по эпизоотологии чумы
Очевидно, что ландшафтно-географические особенности природных очагов определяют видовой состав носителей и переносчиков чумы, а, следовательно, и присущие им особенности эпизоотий. В связи с этим целесообразно остановиться на некоторых вопросах.
Как уже указывалось, принято отличать восприимчивость к инфекции от чувствительности к ней, причём под первой понимают способность заражаться, тогда как вторая характеризует интенсивность, остроту патологического процесса [Олсуфьев Н. Г., Дунаева Т. Н., 1970]. С этих позиций восприимчивость надо считать видовым признаком, а чувствительность индивидуальным.
Многие считают, что одним из главных атрибутов основного носителя должна быть относительная резистентность к чуме. Одним из первых, эту мысль по-видимому, высказал Neter [1948, цит. по Pollitzer R., 1954]. Приверженцы указанной идеи подкрепляют её представлениями о коэволюции носителя и возбудителя чумы [Кучерук В. В., 1965; Pалль Ю. М., 1965]. По их мнению, носителями чумы становились лишь те виды, которые выживали после многократных «встреч с чумой», а в качестве аргументов приводятся данные о меньшей чувствительности к чуме грызунов в эндемичных районах, по сравнению с грызунами в местах, свободных от неё [Ралль Ю. М., 1965; Wu Lien-Teh et al., 1936; R. Pollitzer, 1954; и др.]. В наиболее категоричной форме это сформулировано А. И. Дятловым [1989]: «… эпизоотии чумы — достаточно эффективный, широко распространенный и на отдельных территориях постоянно действующий фактор естественного отбора…».
Однако, по мнению М. П. Козлова [1979], «…устойчивость видов к чуме не обусловлена обитанием их на энзоотичной территории, а степень инфекционной чувствительности и инфекционной восприимчивости грызунов к чуме не может служить показателем их важности в поддержании эпизоотического процесса.»
Очевидно, истина лежит где-то посредине между этими крайностями, так как имеется много фактов свидетельствующих о том, что по чувствительности к чуме популяции носителей неоднородны. Последнее позволило Ю. М. Елкину [1960] разделить грызунов на 4 группы:1) высокочувствительные (домовая мышь, тамарисковая и малоазиатская песчанки, песчанка Виноградова, степная пеструшка, слепушонка); 2) залегающие в спячку с резко меняющейся по сезонам чувствительностью (сурки, суслики); 3) незалегающие в спячку, в популяциях которых всегда имеются высокочувствительные и очень резистентные особи (полуденная, большая, краснохвостая песчанки, серая крыса, обыкновенная и общественная полевки, полевка Брандта, полевая мышь); 4) чувствительные только к очень большим дозам заражения (лесная мышь).
Понятно, что список видов, входящих в каждую из групп, приведенных Ю. М. Елкиным, может быть существенно расширен. Как считал Ю. М. Ралль [1965], «…виновниками укоренения чумы могут стать лишь представители второй и третьей групп с неодинаковой чувствительностью…». Насколько мы можем судить, на аналогичных или близких позициях стоят сейчас и зарубежные ученые. В частности T. Butler [1983] пишет: " Классическая теория резервуаров чумы требует одного или нескольких видов мелких млекопитающих, которые служат в качестве резервуаров «энзоотии», и одного или нескольких относительно чувствительных видов, выступающих в роли «эпизоотических хозяев» и могут вовлекаться в так называемый «rat die-offs " или " ratfalls». В очагах городской чумы одни и те же виды крыс могут иметь значение как «энзоотичные», так и «эпизоотичные» виды, в то время как в сельских районах или районах «дикой» чумы обычно эти функции распределяются между разными видами грызунов. Сказанное иллюстрируется следующими примерами. В Индии основным носителем инфекции является резистентная к чуме песчанка Tatera indica, а звеном между нею и человеком — домовые крысы. На Яве же в роли носителя чумы выступает резистентная к ней полевая крыса R. exulans, а функция передаточного звена сохраняется за домовыми крысами. Однако подобная закономерность пока не подтвердилась для очагов дикой чумы в США. Там наиболее частыми «промежуточными» носителями являются разные виды земляной белки и луговой собачки (Cynomys), однако единого мнения об основном носителе нет. В Нью-Мексико и Аризоне чумной микроб персистирует на койоте, американской рыси и кроликах, но каким образом осуществяется циркуляция микроба между ними, остается загадкой.
У грызунов, как и у людей (см. ниже), описаны различные клинические формы чумы: бубонная, септическая и первичная легочная. Возникновение тех или иных форм связано с механизмом заражения и локализацией входных ворот инфекции, хотя из этого правила есть и исключения. Так, алиментарный путь заражения может приводить к развитию либо бубонной, либо первичной легочной чумы. Все же, поскольку чума относится к числу кровяных инфекций с трансмиссивным механизмом передачи, основными формами её у грызунов надо считать бубонную и септическую.
Интенсивное размножение Y. pestis и связанная с ним интоксикация сопровождается глубокими дистрофическими и некробиотическими изменениями в ретикуло-эндотелиальной системе, приводящими к сепсису и гибели животных. Но процесс может останавливаться на ранних стадиях заболевания, о чём свидетельствуют находки в природе грызунов со следами перенесенной чумы [Лобанов В. Н., Федоров В. Н., 1938; Некипелов Н. В., 1953; Williams J. E., Cavanaugh D. C., 1983]. В других случаях процесс сразу переходит в септическую фазу, при которой патологоанатомических изменений не выявляют.
Помимо острых форм, у грызунов описаны затяжные и хронические формы, значение которых для эпизоотологии оценивается неоднозначно [Ралль Ю. М., 1965; Козлов М. П., 1979; Wu Lien-Teh et al.,1936; Pollitzer R., 1954]. Имеются также сообщения об «инаппарантном» течении чумы, которое при определенных, пока не выясненных, условиях может сопровождаться бактериемией — основным условием трансмиссивной передачи инфекции. Однако, как указывал R. Pollitzer, инаппарантную чуму нельзя смешивать с латентной формой заболевания у грызунов во время спячки.
Рассмотрим теперь особенности течения чумы у основных носителей, указанных в табл. 4, и начнем с сурков и сусликов, имеющих много общего.
В соответствии с образом жизни эпизоотии среди малых сусликов носят характер выраженной зависимости от времени года. Как указывали В. Н. Федоров и соавт. [1955], ранневесенние эпизоотии обычно краткосрочны; продолжительность их равна периоду спаривания, и они очень редко приобретают разлитое течение, хотя затрагивают ослабленных зимней спячкой грызунов. Тем не менее такие эпизоотии играют большую роль в сохранении возбудителя в природных очагах, поскольку сопровождаются обновлением и увеличением численности блох, оставшихся со времени последних эпизоотий. Наступающие вслед за спариванием биологические циклы в жизни грызунов — беременность и лактация — резко уменьшают подвижность (активность) сусликов и обрывают эпизоотии.
Вторая эпизоотическая волна, более постоянная и широкая чем первая, наступает в конце весны — начале лета и совпадает с началом расселения молодняка по пустым заблошивленным норам. Другие причины весенне-летних эпизоотий: а) значительно возросшая плотность популяций грызунов; б) ухудшение физиологического состояния молодняка, часто в результате истощения кормовой базы; в) непрерывное увеличение численности блох.
После завершения расселения молодняка начинается постепенное залегание сусликов в спячку, что приводит к затуханию эпизоотий.
Сурки существенно отличаются от сусликов рядом экологических особенностей, в частности семейным образом жизни: в одной норе, помимо родителей, живут сурчата, родившиеся в этом году, а иногда и сурки прошлого выводка. Естественно, что такой образ жизни накладывает отпечаток на характер эпизоотий, которые очень редко приобретают большие масштабы [Wu Lien-Teh et al., 1936]. Несмотря на это, зараженных чумой сурков и их эктопаразитов можно обнаружить в течение всего периода их бодрствования. Кстати, в отличие от сусликов суркам свойственна только зимняя спячка [Федоров В. Н. и др., 1955]. В эпидемиологическом отношении особенно опасны сурки в период после окончания линьки, когда на них начинается охота ради шкурок. Правда, сейчас промысел сурка почти повсеместно резко сократился.
Говоря о чуме у сурков, надо отметить, что в переносе возбудителя особо важное значение имеют их блохи — O. silantiewi. Эти блохи хорошо приспособились к низким температурам в норах сурков и потому служат длительными хранителями возбудителя [Иофф И. Г., 1941].
Для сусликов и сурков характерно латентное течение инфекции во время зимней спячки. У тарбаганов это было доказано еще в 20-х годах Wu Lien-Teh и R. Pollitzer [цит. по Wu Lien-Teh et al., 1936]. Исследователи установили, в частности, что во время спячки сурков чумной микроб сохраняется без потери вирулентности на месте введения и/или в регионарных лимфатических узлах. Через некоторое время после пробуждения у животных может развиваться острый процесс, сопровождающийся бактериемией. Однако нечувствительность спящих сурков к чуме не является абсолютной, поскольку они все-таки могут погибать от инфекции, а иногда на коже спящих сурков в местах заражения развиваются язвы, инфицированные чумным микробом, что не свойственно бодрствующим животным [Ралль Ю. М., 1965].
Латентная инфекция у сурков становится явной весной, после пробуждения. Аналогичная картина наблюдается у сусликов: латентное течение во время спячки с генерализацией процесса и гибелью отдельных зверьков после пробуждения [Гайский Н. А., 1925].
Описание чумы у сурков и сусликов будет неполным, если не упомянуть еще одну особенность патогенеза, а именно вариабельность процессов в активные периоды жизни грызунов. Так, описывая одну из эпизоотий, И. С. Тинкер [1940] указывал, что в ходе первого месяца (июль) эпизоотии смертность сусликов была очень высокой и обнаружение их трупов в степи в состоянии сильного окоченения были обычным явлением. Однако ни бубонов, ни видимых изменений во внутренних органах при этом у них не находили, хотя в отпечатках из органов обнаруживалось огромное число микробов. К середине лета у сусликов стали чаще обнаруживаться крупные бубоны (размерами до лесного ореха), увеличенная селезенка и некротические изменения в печени, но выделить возбудителя у таких сусликов было трудно, а в некоторых случаях и невозможно.
Существенные различия в патологоанатомической картине от весны к осени отмечаются также у больных сурков. Весной чаще встречаются остросептические формы заболевания с большим числом бактерий во всех органах и крови, а к концу лета нарастают продуктивные изменения и резко падает вероятность выделения чумного микроба [Петрунина О. М, 1951а].
Иная ситуация наблюдается у песчанок.
Полуденные песчанки активны круглый год. В соответствии с этим зараженных зверьков и их блох обнаруживают постоянно, но все же чаще в весенне-летний и осенне-зимний сезоны. Подобную закономерность, т. е. двугорбый характер эпизоотий, В. Н. Федоров и соавт. [1955] связывали с сезонными изменениями активности полуденных песчанок, возможными изменениями их физиологического состояния, а также с влиянием сезонных условий на видовой состав и активность блох, паразитирующих на грызунах. В известной мере это мнение основывалось на экспериментальных данных, опубликованных В. Н. Лобановым и В. Н. Федоровым еще в 1938 г. Согласно им, в период с июля по октябрь большинство полуденных песчанок погибало от острых генерализованных форм чумы, а с апреля по июль у них отмечались локализованные формы с явной тенденцией к разрешению процесса, причём выделить культуру Y. pestis от таких животных не удавалось даже через биопробы.
Необходимо напомнить, что полуденные песчанки являются носителями чумы в песчаном волго-уральском очаге, тогда как на правом берегу Волги, в степном очаге северо-западного Прикаспия, в поддержании чумы они никакой роли не играют [Ралль Ю. М., 1965]. Объяснение столь парадоксальной ситуации было найдено Е. С. Бирюковой [1960], которая впервые в 1955–1957 гг. подметила различия в чувствительности к чуме полуденных песчанок, обитающих на левом и правом берегах реки Волги: чувствительность к чуме первых оказалась на несколько порядков выше, чем вторых. При этом очень важно отметить, что эти различия чувствительности двух популяций полуденных песчанок выявлялись не только при искусственном заражении, но и в случае укусов их инфицированными блохами [Ю. М. Елкин, 1960].
Вопросами чумы у полуденных песчанок много занимались М. И Леви и его сотрудники, подтвердившие первоначальное наблюдение Е. С. Бирюковой (табл. 6). При этом были установлены существенные различия в патогенезе чумы у право- и левобережных песчанок, подтверждающие гипотезу о том, что основным носителем чумной инфекции в природе являются «не столько высоко восприимчивые к чуме, сколько малочувствительные грызуны с высоким уровнем бактериемии». Кроме того, М. И. Леви, одному из немногих, удалось показать, что «резистентность к чумному микробу передается по наследству» (табл. 7). Небезынтересно, что отмеченные выше различия в чувствительности полуденных песчанок к чуме не распространяются на их чувствительность к псевдотуберкулёзу, туляремии и бруцеллезу, т. е. достаточно специфичны.


Не меньший интерес представляют сведения о заболеваемости чумой больших песчанок, обитающих на огромных просторах равнинного среднеазиатского очага. Здесь сохраняется в общем тот же характер эпизоотий, как и в волго-уральском очаге. Зараженных чумой больших песчанок можно обнаружить в любой сезон, но пики эпизоотий также приходятся на весну и осень. Зимние спады объясняют резким уменьшением активности песчанок, тогда как подъемы связывают с повышением подвижности грызунов и увеличением численности блох рода Xenopsylla. Однако число зараженных особей в популяциях песчанок в различные сезоны бывает неодинаковым, что опять-таки объясняется неодинаковой подвижностью грызунов в различное время года. Обращают на себя внимание отличия в инфекционной чувствительности отдельных возрастных групп, разброс индивидуальной чувствительности и зависимость её от сезона. Зарегистрированы и различия чувствительности к чуме у больших песчанок в разных популяциях. Например, зверьки из северо-восточного Прикаспия по сравнению с песчанками из Туркмении и особенно из Или-Каратальского междуречья оказались более чувствительными к инфекции [Наумов Н. П. и др., 1972]. Интересно, что в период затухания эпизоотий увеличивается процент затяжных и хронических форм чумы, столь характерных для больших песчанок. При этом часто выделяются авирулентные штаммы чумного микроба. Относительно вялое течение чумы с тенденцией к переходу в хронические формы подтверждено в экспериментах О. М. Петруниной [1951б] и А. А. Левиной [1960]. В опытах последней некоторые песчанки жили по несколько месяцев после заражения, но септицемия у них отмечалась очень редко, да и то в тех случаях, когда они погибали не позднее 35 сут.
Относительная устойчивость к чуме наряду с индивидуальными колебаниями чувствительности и тенденцией к затяжному течению присуща также краснохвостой и некоторым другим видам песчанок [Ралль Ю. М., 1965]. В отличие от них такие песчанки, как тамарисковая и Виноградова или Tatera brantsi, обладают высокой чувствительностью, на которую мало влияют климатические и другие факторы. В частности, тамарисковая песчанка легко вовлекается в эпизоотии среди грызунов иных видов и быстро погибает от острых септических форм чумы, что не раз позволяло использовать их в качестве хороших объектов для биологических проб на чуму.
В отличие от зимоспящих грызунов и песчанок представители семейства заячьих — пищухи стали объектами внимания относительно недавно.
В ходе эпизоотий среди сурков и сусликов в Забайкалье часто находили трупы пищух, павших от чумы; при вскрытии у них обнаруживали картину сепсиса. Это побудило Н. А. Гайского и Н. Д. Алтареву [1944] провести соответствующие исследования. Используя разные заражающие дозы, они установили, что от чумы погибает около 50 % даурских пищух. Тем не менее их все еще продолжали относить к числу второстепенных носителей [Некипелов Н. В., 1959б]. Однако отношение к пищухам изменилось, когда в 1955 г. в относительно спокойном районе Северо-Западной Монголии, недалеко от границы с Горным Алтаем, среди пищух была зарегистрирована эпизоотия чумы, а в 1961 г. она появилась и здесь. Сначала эпизоотию среди монгольских пищух в Горном Алтае рассматривали как результат заноса чумы из Монголии [Домарадский И. В. и др., 1963а], но в последующем очаг чумы на северном и южном склонах хребта Сайлюгем стали относить к издавна существующим очагам северо-восточной окраины центрально-азиатской «чумной зоны» [Голубинский Е. П. и др., 1987].
Очаг в Сайлюгеме полигостальный, но роль основного носителя в нем играет монгольская пищуха, а дополнительными носителями являются даурская пищуха, длиннохвостый суслик и плоскочерепная полевка. Кроме того, в эпизоотии иногда вовлекаются алтайский сурок, тушканчик-прыгун и даурский хомячок. Функции перносчиков выполняют по крайней мере 5 видов блох, ни один из которых самостоятельного значения не имеет.
Эпизоотии в Сайлюгеме начинаются весной и достигают пика, совпадающего по времени с выходом на поверхность и расселением молодняка, в июле-августе. Затем начинается постепенный спад, но эпизоотии не прекращаются даже зимой [Логачев А. И., Михайлов Е. П., 1984].
Ю. М. Ралль [1965] считал, что пищухи относятся к числу высокоустойчивых видов. При этом он ссылался на упоминавшиеся уже опыты Н. А. Гайского и Н. Д. Алтаревой. Однако новые исследования заставили пересмотреть подобное мнение. Основанием послужили прежде всего данные В. Е. Тарасовой и А. М. Шамовой [1966] о том, что пищухи весьма чувствительны к чуме, если она вызывается штаммами чумного микроба из сайлюгемского очага. Как подчеркивали Е. П. Голубинский и соавт. [1987], это оказалось необычным и принципиально новым фактом, так как он идёт вразрез с постулатом, согласно которому высокочувствительные к чуме животные не могут быть носителями чумы. Небезынтересно добавить, что пищухи чувствительны даже к авирулентным для других животных штаммам чумного микроба, включая вакцинный штамм EV [Иннокентьева Т. И., 1969], хотя по реакции на эти штаммы пищухи из Тувы несколько отличаются от горно-алтайских [Тарасова В. Е., Иннокентьева Т. И., 1971].
В заключение остановимся на особенностях течения чумы у крыс, которых сейчас насчитывается около 500 видов. Большинство крыс обитают в диком состоянии и только два вида — черная (R. rattus) и серая (R. norvegicus) — приспособились к условиям жизнедеятельности человека и расселились почти по всем материкам [Козлов М. П., 1979]. Неудивительно поэтому, что именно этим видам принадлежит главная роль в распространении чумы среди людей [Wu Lien-Teh et. al., 1936]. Очень точно «крысиная» чума описана в романе Альбера Камю[8].
Течение чумы у крыс в естественных условиях хорошо известно еще со времени работы Индийской комиссии в период с 1906 по 1917 г., опубликовавшей большую серию отчетов. Подробно описано оно также русскими исследователями во время одной из последних, но наиболее значительных вспышек бубонной чумы в Одессе в начале века.
Обычно эпизоотии среди крыс, на фоне их высокой численности, предшествуют заболеваниям людей. При этом трупы погибших животных часто находят на улицах, поскольку в состоянии агонии крысы покидают свои норы и «…бегут, пошатываясь, прямо вперед, падают в судорогах и умирают при дневном свете…» [Кияницын И. И., 1911]. При внешнем осмотре погибших крыс часто обращает на себя внимание сильно выраженное трупное окоченение, которое сохраняется даже у загнивших трупов [Wu Lien-Teh et al., 1936].
Как правило, от больных крыс человек заражается блохами, покидающими трупы животных. Иные механизмы передачи инфекции существенного значения не имеют [Pollitzer, R., 1954].
При вскрытии трупов крыс самым ярким признаком чумы являются бубоны, в 75 % случаев цервикальные. На разрезе бубоны имеют типичный вид вследствие некрозов сердцевинной части. Кроме того, отмечается резкое увеличение других лимфатических узлов. Очень характерным для крыс можно считать гиперемию подкожных сосудов, что придает красноватый оттенок коже, особенно заметный на подошвах задних лап, а также на поверхностных мышцах груди и живота. Обнаруживаются геморрагии вокруг бубонов. В печени — все стадии жировой дистрофии. В начальной стадии заболевания контраст между пораженными участками, жёлтоватого цвета, и здоровыми, красноватыми, придает этому органу вид мрамора. Позднее вид её становится более однородным, и почти у всех крыс из-за очагов некроза печень как бы «посыпанна перцем» [Кияницын И. И., 1911].
Патологоанатомические изменения в других органах и тканях крыс весьма сходны с таковыми у людей, погибших от чумы (см. раздел 4.2).
Вскрытие крыс, впрочем как и других грызунов, дает возможность взять материал для бактериологического исследования, однако в случае разложившихся трупов рекомендуется прибегать к постановке биопроб с применением «австрийского» метода заражения морских свинок.
Помимо острых форм чумы для крыс особенно характерны хронические формы, при которых выявляются творожистое перерождение бронхиальных желёз, индурация ткани легких или некротические очаги в селезенке. Важно, что такие случаи могут сопровождаться бактериемией [Wu Lien-Teh et al., 1936]. Встречаются также инаппарантные формы, когда нет ни макроскопических изменений во внутренних органах, ни бактериемии, но возбудитель удается обнаружить с помощью биопроб. Однако мнения исследователей о роли таких форм в поддержании и распространении чумы далеко не единодушны [Pollitzer R., 1954].
Картина заболевания крыс при экспериментальном заражении в общем не отличается от описанной выше [Анисимова Т. И., 1962].
Как чёрные, так и серые крысы весьма чувствительны к чуме, но в естественных условиях серые заражаются легче, чем чёрные. В Индии, например, были случаи, когда эпизоотии сначала начинались среди серых крыс и лишь позднее охватывали чёрных. В то же время по чувствительности к чуме популяции крыс столь же неоднородны, как и популяции многих других животных. Так, по данным Т. И. Анисимовой (1962), наряду с особями, не чувствительными к заражению 25-100 млн. клеток Y. pestis, попадаются такие, которые погибают от единичных клеток. Однако в тех же опытах наблюдалась зависимость чувствительности к чуме обоих видов крыс от возраста животных, но сезонных изменений её не выявлено. Видимо, поэтому эпизоотии среди крыс встречаются в любое время года, в частности в Индии [Кашкадамов В., 1901] и других странах тропического пояса [Butler T., 1983]. Вместе с тем нельзя не указать на неодинаковую чувствительность к чуме крыс из эндемичных и свободных от неё областей.
В заключение раздела приведу цитату из работы М. П. Козлова [1979]: «… синантропные и полусинантропные крысы как весьма пластичные виды являются процветающими. При снижении контроля за численностью их эктопаразитов они могут вновь стать одним из основных потенциально опасных видов грызунов — источников чумы в тропических и субтропических районах…». И следует добавить, что типичные крысиные очаги чумы все еще остаются слабо изученными [Комитет экспертов ВОЗ по чуме, 1971] и таят в себе много неожиданностей.
2.4. Эпидемиология
Несмотря на то, что от начала эпидемии чумы в Гонконге нас отделяет 100 лет и за это время мир стал неузнаваемым, чума не перестала быть грозной. Вспышки чумы продолжают регистрироваться ежегодно (см. табл. 1 и 2), причем источником их остаются «старые» природные очаги в Африке, Азии и Америке, только изменился их удельный вес. К примеру, случаи чумы теперь редки в Индии или Китае, но много хлопот начали доставлять очаги в Юго-Восточной Азии и в Америке. По данным ВОЗ, за 10 мес. 1996 г. в мире было зарегистрировано 234 случая чумы (1 — в Монголии, 23 — в Перу, 2 — в США и 208 — на Мадагаскаре).
К сожалению, многие особенности эпидемиологии чумы на территории бывшего Советского Союза остаются для нас тайной за семью печатями, хотя, как показано в табл. 3, только за 60 лет в нем было зарегистрировано около 4 тыс. заболеваний!. Поэтому при эпидемиологическом анализе приходится апеллировать к дореволюционным работам или исследованиям зарубежных учёных и картина, конечно, оказывается неполной. Все сказанное оставляет чувство большой неудовлетворенности и вот почему.
Все эпидемии чумы в России в XIX столетии и до установления советской власти условно можно разделить на две категории. К первой мы относим эпидемию чумы во Ветлянке и вокруг неё (сентябрь 1878 г. — январь 1879 г.), т. е. эпидемию, которая возникла почти за 15 лет до начала последней пандемии чумы (1894 г.). Истинная причина её осталась неразгаданной. Однако расположение Ветлянки на Волге, в местах повсеместных поселений малого суслика — одного из основных резервуаров чумной инфекции, позволяет нам с большой долей вероятности говорить о его причастности к этой эпидемиии. Весьма вероятно, что с эпизоотиями на малом суслике была также связана вспышка чумы в селе Колобовка на левом берегу реки Ахтубы в Астраханской губернии (июль — август 1899 г.), хотя Д. Д. Ахшарумов (1900) не исключал возможности её заноса паломниками из Северо-Восточной Монголии. К этой же категории «аутохтонной» чумы мы причисляем эпидемии «тарбаганей болезни» в Забайкалье (1910–1916 гг.).
Ко второй категории, по нашему мнению, принадлежат эпидемии чумы в Одессе, включая последнюю вспышку 1910 г. Все они протекали по типу «городской чумы», т. е. носителями возбудителя были крысы. Поскольку природных очагов чумы на Украине нет, происхождение чумы в Одессе можно объяснять только заносами извне, с чем в общем все согласны.
Сопоставление приведенных фактов показывает, что серьезные открытия делались только тогда, когда причину чумы искали «дома». Ведь именно благодаря такой направленности поиска были заложены основы учения о природной очаговости инфекций Д. К. Заболотным и его последователями, в частности И. А. Деминским поплатившимся за это жизнью.
Возвращаясь к сегодняшнему дню, приходится констатировать, что под спудом находится или утрачен колоссальный материал, касающийся особенностей проявления чумы у многих сотен людей, проживавших на огромной территории, которые занимают её природные очаги. Это особенно досадно, так как материалы собирались большой армией чумологов, равной которой нет в мире.
Так как чума является типичным зоонозом, её вспышкам у людей всегда предшествуют эпизоотии. Первые случаи заражения человека чумой от больных грызунов возникают в результате укусов блох или прямого контакта с животными. При этом заболевания людей носят ясно выраженный зоонозный характер и клинически начинаются как локальные или внутренне-диссеминированные формы (по классификации Г. П. Руднева, 1938), при которых больные практически не заразны. Основным источником инфекции человек становится в тех случаях, когда указанные формы чумы осложняются вторичной пневмонией. Тогда чума начинает распространяться по типу капельных инфекций и становится особенно опасной. Однако заразными для окружающих являются также моча и испражнения больных. Специальные меры безопасности должны соблюдаться при вскрытии и захоронении трупов погибших [Лобанов В. Н., 1956].
Почему вторичные пневмонии не всегда и не везде дают начало эпидемиям легочной чумы? На этот вопрос до сих пор ответа нет. Известно только, что их возникновение не связано с какой-то особой вирулентностью чумного микроба [Wu Lien-Teh et al., 1936; Girard G., 1963] или его гипотетической «пневмотропностью» [Pollitzer R., 1954]. Известно также, что вспышки легочной чумы наблюдаются преимущественно в природных очагах умеренного климата; в очагах тропического пояса они носят эксквизитный характер. К числу же способствующих факторов часто относят переохлаждение, перенаселение городов, антисанитарные условия, нищету.
Как уже указывалось, при естественном заражении обычно развивается одна из форм бубонной чумы, но описаны случаи, когда бубоны не выявляются и заболевание начинается как первичные пневмония, сепсис и реже менингит [Butler T., 1983]. Именно поэтому для окружающих опасен любой больной чумой.
Главная роль среди переносчиков чумной инфекции принадлежит блохам; другие кровососущие членистоногие — вши, постельные клопы, клещи и летающие насекомые, если и играют роль, то второстепенную (см. выше). Чаще всего блохи заражают людей в полевых условиях (в природных очагах) и в населенных пунктах во время эпизоотий на крысах. Тут однако надо обратить внимание на следующий факт. Судя по данным американских ученых [Mann J. M. et al.,1979], хорошо изученные случаи заболевания чумой были связаны не столько с работой или пребыванием в полевых условиях, сколько с появлением грызунов в жилищах человека. Эти наблюдения легко увязываются с предположением C. V. Reyn и соавт. [1977] о том, что домашние собаки и кошки служат звеном в передаче инфицированных блох от грызунов к человеку. Не этим ли объясняется факт, подмеченный еще старыми авторами, что чума «проникала далеко не во все дома и семьи» [Ахшарумов Д. Д., 1900]? Кроме того, не следует забывать об указании на передачу инфекции блохами от человека к человеку [Наркевич М. И. и др., 1991] и на заражение при раздавливании насекомых зубами [Butler T., 1983].
Немалое эпидемическое значение для возникновения чумы среди людей имеют такие домашние животные, как верблюды, что мало известно на Западе, а также козы [Christie A. B. et al.,1980] и дикие кролики, о чём не знают в нашей стране (табл. 8 и 9). В подобных случаях люди заражаются главным образом при разделке туш больных животных и употреблении их мяса в пищу. По-прежнему, хотя и в меньших масштабах, заражение бывает следствием охоты на сурков и сусликов. В таких случаях заболевания у людей протекают очень тяжело и часто оканчиваются смертью больных.


Восприимчивость человека к чуме очень высока, хотя при прочих равных условиях ей особенно подвержены дети и лица, моложе 20 лет. По данным T. Butler [1983], среди заболевших чумой в период между 1925 и 1975 г. на эту категорию приходилось 65 %. Частоту заболеваемости в зависимости от пола определить трудно. Так, в США за указанный период взрослые мужчины заражались примерно в 2 раза чаще, чем женщины, но среди детей такой разницы не было, а к 80-м годам разница в частоте заболеваемости женщин и мужчин нивелировалась. Однако во время эпидемии чумы в Ветлянке «влияние пола осталось незаметным, влияние же возраста выяснилось отчасти: с возрастанием годов прогрессирует и число смертности; детский возраст менее расположен к заболеванию» [Ахшарумов Д. Д., 1900]. Скорее всего, группы риска определяются обычаями и социально-экономическими условиями жизни населения. То же можно сказать и об этнических особенностях.
Повторных заболеваний чумой не бывает, хотя отдельные случаи их описаны [Афанасьев М. И., Вакс П. Б.,1904; Butler T., Hudson W. B., 1977]. Данные последних авторов позволяет высказать предположение, что современная терапия не способствует развитию постинфекционного иммунитета, Как тут не вспомнить пророческие слова Н. Н. Жукова-Вережникова [1940] о том, что основным в лечении чумы должно быть стремление удлинить «сроки жизни больного до предела, за которым начинает проявляться активный иммунитет организма»?
Говоря об эпидемиологии чумы, нельзя обойти молчанием вопрос о её сезонности и периодичности. Как подчеркивал Комитет экспертов ВОЗ по чуме [1971], «…эпизоотический процесс в каждом природном очаге имеет свои особенности с точки зрения цикличности и периодичности. Различные биологические и абиотические факторы определяют экологию местных грызунов — резервуаров инфекции и блох — переносчиков и вызывают сезонные, четко выраженные различия эпизоотологии чумы с характерными пиками активности при благоприятствующих условиях. К сожалению, нет данных, достаточных для того, чтобы связать все это с одним каким-то фактором». Этими вопросами много занимались сотрудники противочумной службы России, хотя немало было сделано и другими исследователями [Wu Lien-Teh et al., 1936; Pollitzer R., 1954, 1960; Bahmanyar M., Cavanaugh D. C., 1976; и др.]. Поэтому, несмотря на пессимистическую нотку в приведенной выше цитате, некоторые обобщения, по крайней мере в отношении сезонности, сделать все же можно.
Зимоспящие грызуны (сурки, суслики) проводят на поверхности земли только 5–7 мес. Из-за повышенной устойчивости к чуме осенью перед залеганием в спячку, а также после пробуждения весной эпизоотические процессы среди них протекают вяло. Острые эпизоотии, как правило, возникают, когда грызуны покидают норы и когда начинается расселение молодняка по новым норам. Именно это сравнительно короткое время года является наиболее опасным для людей. У незимоспящих грызунов и зайцеобразных (песчанок, полёвок, пищух, морских свинок, кроликов и др.) интенсивные эпизоотии продолжаются значительно дольше и часто имеют два пика, совпадающие с периодами массового размножения животных. Высокая миграционная активность блох многих видов носителей, например песчанок, в сочетании с их способностью питаться на других животных и человеке приводит к иррадиации инфекции и более частым вспышкам среди людей. О возможной причине сезонности эпизоотий среди крыс в широтах умеренного климата говорилось выше. В тропической зоне имеются свои закономерности, связанные с географическим положением очагов «крысиной чумы» и эколого-физиологическими особенностями животных.
Необходимо однако отметить, что на сезонность и периодичность эпизоотий чумы, а, следовательно, и на её эпидемиологию существенное влияние может оказывать деятельность человека. Яркой иллюстрацией служат интенсивные эпидемии чумы во Вьетнаме в 60-х годах, одной из причин которой были изменения в экологии грызунов и их блох, обусловленные широким применением дефолиантов в природных биотопах [Акиев А. К. и др., 1983].
Помимо сезонной периодичности вспышек чумы, отмечены и другие её особенности. Так, в штате Шолапур (Индия) пик заболеваемости отмечается раз в 5 лет, в Бразилии зарегистрированы 5- или 10-летние циклы, в Аргентине 4-летние и в Южной Африке — 5-6-летние циклы [Pollitzer R., 1954]. В этом контексте небезынтересно указать на наличие определенной связи между эпидемиями чумы и солнечной активностью. Установлено, что существует цикличность солнечной активности, максимумы которой отделены друг от друга 11-летними периодами с колебаниями от 7 до 17 лет. Используя многочисленные литературные данные,А. Л. Чижевский [1973] подметил, что свыше 60 % всех известных эпидемий чумы (VI–XVII вв.) «…хорошо совпадают с датами эпох солнечных максимумов». Правда, начиная с XVIII в. эта закономерность нарушается. Причиной этого, по мнению А. Л. Чижевского, кроется в том, что история донесла до нас только наиболее крупные эпидемии, сравнимые с другими земными катаклизмами, а в XVIII в. и позднее таких эпидемий не было. Каким образом солнечная активность связана с возникновением эпидемий чумы (а также холеры, гриппа, дифтерии) предстоит еще выяснить. Может быть солнечная активность как-то влияет на возбудителя, а возможно она прямо или косвенно способствует размножению грызунов — носителей чумы. Некоторые соображения по этому поводу высказал М. С. Эйгенсон [1957]. Неисключено, однако, что большее значение, нежели солнечная активность, во всех этих процессах имеет ионизирующее излучение, связанное со вспышками сверхновых звезд [Бяков В. М., 1996]. Так или иначе, но дальнейшее изучение проблем цикличности чумы может способствовать составлению долгосрочных прогнозов, наличие которых необходимо для эффективной борьбы с этой инфекцией.
В заключение остановимся еще на двух важных вопросах. Прежде всего, мы имеем в виду бактерионосительство. О его возможном значении при чуме говорили многие исследователи, начиная с Е. В. Падлевского [1915]. Однако вполне очевидным оно стало лишь после работы J. D. Marshall и соавт. [1967], которые выделили Y. pestis из зева у 15 из 114 здоровых людей; при отсутствии химиопрофилактики бактерионосительство продолжалось до 31–35 дней. Второй вопрос касается возможности заражения грызунов от больных людей, которую допускал, в частности, Б. К. Фенюк [1959]. Соглашаясь с ним, Ю. М. Ралль [1965] подчеркивал тем не менее, что «внутридомовая» чума укореняться не может. В данном случае он был, по-видимому, солидарен с R. Pollitzer [1954], упоминавшим лишь о редких случаях заражения крыс во время эпидемий легочной чумы в Харбине (1910–1911) и во Владивостоке (1920–1921), не имевших серьезных последствий. В то же время нельзя не считаться с возможностью заражения блох от людей больных чумой, поскольку блохи могут служить источником новых заболеваний людей без участия грызунов (см раздел 2.2). При этом опасны как X. cheopis, так и P. irritans [Новикова Е. И., Лалазаров Г. А., 1944; Blanc G., Baltazard M., 1943]. Поэтому тезис о «незаразности» больного бубонной чумой не следует принимать безоговорочно.
До сих пор основной акцент мы делали на вспышках чумы, которые связаны с пребыванием людей в эндемичных очагах. Однако не следует забывать, что чума угрожает им и в странах, которые расположены далеко от этих очагов. Источником инфекции в таких случаях могут быть лица, заразившиеся в природных очагах и прибывающие в страну еще без явных признаков заболевания. Другим источником инфекции служат крысы, особенно гнездящиеся в контейнерах, где их трудно обнаружить при санитарно-карантинных досмотрах судов. При этом надо опасаться не только непосредственного контакта с больными грызунами, но также их блох, из числа которых наибольшее значение имеют виды, указанные в табл. 10. Как подчеркивали M. Bahmanyar и D. C. Cavanaugh [1976], перечисленные виды блох отличаются двумя важнейшими особенностями. Во-первых, они легко инфицируются чумным микробом и предрасположены к блокообразованию, а во-вторых, они тесно связаны с человеком, нападают на него, когда голодают, и легко адаптируются к городским условиям.

Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги