История мембранной теории биопотенциалов
Главенствующая и единственная на сегодняшний день теория распространения нервного импульса заслуживает отдельной истории.
Она интересна во многих аспектах: и как победившая идеология, и как очень элегантная, даже изящная, или изощрённая, логическая модель. Также интересна эта история полным отсутствием альтернативных идей и, следовательно, она не выверена в спорах с оппонентами. Сплошные единомышленники. Вспоминается один из абсурдных советских лозунгов: «Учение Ленина истинно потому, что оно верно!». И продолжая ленинскую терминологию, давайте рассмотрим три источника, три составные части мембранной теории – осмос, теорию электролитической диссоциации и гипотезу Бернштейна.
Осмос
Началом этой истории послужило открытие осмоса. Думаю, что не все наши читатели слышали это слово раньше, так вот, осмос – это процесс, в котором при определённом давлении растворитель проходит через полупроницаемую мембрану из более концентрированного в менее концентрированный раствор. При этом мембрана пропускает растворитель, но не пропускает растворённые в нём вещества.
Впервые в 1748 году осмос наблюдал Жан-Антуан Нолле (Jean-Antoine Nollet, 1700—1770) – французский священник (аббат) и физик, член Парижской академии наук с 1739 года.
Легенда гласит, что однажды аббат Нолле слил недопитое им вино во фляжку из свиного мочевого пузыря и погрузил его охладить в бочку с водой. Наутро вынув пузырь из бочки, он обратил внимание, что тот раздулся. Попробовав вино, аббат понял, что напиток стал менее крепким. Нолле сделал вывод, что вино стало разбавленным из-за того, что вода проникла в него сквозь стенку сосуда под действием некой силы, которая возникла из-за разницы концентрации воды в бочке и в вине. При этом аббат заметил, что через стенку пузыря проникла только вода. Если ли бы пузырь с вином, который Нолле положил в воду, не обладал способностью растягиваться, проникающая вода подняла бы давление и процесс бы остановился. Давление, которое необходимо приложить, чтобы не пустить воду в вино называется осмотическим. Оно зависит от разности концентраций растворённых веществ по обе стороны мембраны.
Пример осмоса: к яичной скорлупе с внутренней стороны прилегает плёнка – это тоже полупроницаемая мембрана, она пропускает молекулы воды и задерживает молекулы сахара. Если такой мембраной разделить растворы сахара с концентрацией 5 и 10% соответственно, то через неё в обоих направлениях будут проходить только молекулы воды. В результате в более разбавленном растворе концентрация сахара повысится, а в более концентрированном, наоборот, понизится. При этом объёмы разделённых мембраной растворов изменятся. Когда концентрация сахара в обоих растворах станет одинаковой, наступит равновесие.
Однако целенаправленное исследование этого явления было начато лишь спустя столетие.
В середине XIX века осмотическую проницаемость клеточных мембран изучали физиолог Моль и ботаник Негели. А с конца XIX века к ним присоединились физики и химики, одним из которых был немецкий учёный Адольф Фик. В 1855 году исследуя перенос через искусственные мембраны из нитроцеллюлозы, он сформулировал феноменологические законы диффузии, названные впоследствии его именем.
Одним из выдающихся учёных, работавших в этой области, был Вильгельм Пфеффер (нем. Wilhelm Friedrich Philipp Pfeffer, 1845 – 1920) – немецкий ботаник и физиолог растений. Он изучал осмотические явления, обусловливающие поглощение растениями воды и минеральных веществ. Работы Пфеффера заложили основы мембранной теории клеточной проницаемости.
В 1877 году он изготовил искусственную полупроницаемую мембрану. Для этого в пористый фарфоровый сосуд он налил раствор медного купороса и поместил его в другой сосуд, заполненный раствором ферроцианида калия. В порах первого фарфорового сосуда растворы контактировали и взаимодействовали друг с другом. В результате в них образовывалась плёнка из ферроцианида меди, которая обладала полупроницаемостью.
Далее, полученный сосуд, в порах которого образовалась полупроницаемая мембрана, заполненный раствором сахарозы, помещали в воду.
Такой прибор получил название осмометра. Схема осмометра Пфеффера на Рис.29: 1 – сосуд с растворителем; 2 – мембрана; 3 – ячейка с раствором; 4 – манометр.
В результате своих исследований, Пфеффер установил, что поступление воды в раствор через полупроницаемую перегородку обусловлено разностью концентраций растворов.
Однако Пфеффер не обнаружил какой-либо количественной зависимости осмотического давления от концентрации и температуры. Данную задачу спустя двадцать лет разрешил Вант-Гофф (Jacobus Henricus (Henry) van't Hoff; 1852 – 1911).
В 70-х годах 19 века молодой голландский ботаник X. Де Фриз исследовал влияние осмоса на изменение объёма клеток растений в растворах разной концентрации.
В 1894 году Де Фриз рассказал о работах Пфеффера молодому химику Я. Вант-Гоффу. Внимательно изучив исследования Пфеффера, Вант-Гофф обратил внимание, что осмотическое давление в разных растворах получается одинаковым, если измерять концентрацию не в граммах на литр, а в молях, т. е. существенной является не масса, а число молекул растворённого вещества. Допустив, что молекулы растворённого вещества ведут себя как молекулы идеального газа он решил, что для выражения осмотического давления можно использовать уравнение Менделеева-Клапейрона. [7]

Рисунок 29. Осмометр Пфеффера.
За теорию растворов Вант-Гофф спустя 15 лет получил Нобелевскую премию по химии. Вот такой важный вклад в науку внёс Де Фриз, поговорив с Вант-Гоффом.
Осмос играет исключительно важную роль в живой природе. Это явление лежит в основе корневой системы питания растений. Благодаря осмосу влага в растениях поднимается и удерживается на высоте в десятки метров.
Питание представителей царства животных также происходит благодаря осмосу. Кровь и лимфа животных – это растворы органических и неорганических веществ. Если концентрация веществ в пищеварительном тракте ниже, чем в крови, влага с питательными веществами всасывается в кровь. Если же концентрация веществ в крови по каким-то причинам окажется ниже, чем в кишечнике (животное объелось соли), всасывание меняет направление, организм обезвоживается, животное погибает.
Осмос оказался причиной внутреннего давления в клетке, именно благодаря ему наши клетки выглядят округлыми и упругими.
Растворы, имеющие одинаковое осмотическое давление, называются изотоническими. Если два раствора имеют различное осмотическое давление, то раствор с бо́льшим осмотическим давлением является гипертоническим, а с меньшим – гипотоническим. При нахождении клеток в изотоническом растворе они сохраняют свой размер и нормально функционируют.
Если же поместить клетки в гипотонический раствор, вода из менее концентрированного внешнего раствора станет переходить внутрь клеток, что приведёт к их набуханию, некоторое время клетка ещё может сохранять целостность, но если процесс не прекращается клеточная оболочка разорвётся и её содержимое вытечет наружу. Такое разрушение клеток называется лизисом.
При помещении клеток в гипертонический раствор вода из клеток уходит в более концентрированный раствор, и наблюдается сморщивание (обезвоживание) клеток. Это явление называется плазмолизом.
Живая клетка представляет собой осмотическую систему. Её мембрана хорошо проницаема как для воды, так и для растворённых питательных веществ.
Осмос, в свою очередь, это результат диффузии воды или другого растворителя через полупроницаемую перепонку, вызванной разностью концентраций или разностью химических потенциалов. Наблюдения за данными явлениями позволяют изучить многие свойства клетки.
Теория электролитической диссоциации
Второй основополагающей предпосылкой для современного объяснения механизма возникновения и проведения нервного возбуждения стала теория электролитической диссоциации шведского учёного Сванте Аррениуса (Arrhenius, Svante August, 1859—1927).
Теория Вант-Гоффа отлично подтверждалась для многих растворов, например, для сахарозы или для водного раствора CO2. Но для некоторых веществ осмотическое давление оказывалось вдвое больше расчётного. Погрешность составляла ровно 100%. Вряд ли её можно было объяснить неточностью измерений.
Обдумывая возможные причины этого расхождения, единомышленник Вант-Гоффа Аррениус догадался, что если, например, для поваренной соли давление оказывается вдвое больше расчётного, то значит, в растворе вдвое больше частиц, чем молекул NaCl, то е. молекула NaCl в воде распадается на две частицы: Na и Сl.
Таким образом, суть теории Аррениуса состоит в следующем: при растворении молекул неорганических и органических кислот, гидроксидов и солей они распадаются (дисоциируют) на ионы:
HСl на Н+ и Cl —,
NaOH на Na+ и OH—,
K2SO4 на 2K+ и SO4—.
Ионы представляют собой заряженные частицы, которые состоят из отдельных атомов, или из групп атомов. Именно эти ионы являются носителями электричества в жидкостях, в отличие от металлов, где перенос электричества осуществляют электроны.
Аррениус пришёл к идее электролитической диссоциации. Суть её в том, что частицы, на которые распадаются многие вещества в растворах, и есть те самые ионы – носители электрических зарядов, с помощью которых ещё Фарадей объяснял законы электролиза.
До Аррениуса учёные полагали, что ионы возникают под влиянием электрического тока, но исследования явления осмоса показало, что это не так. Уже в самом растворе даже в отсутствие электричества имеются и движутся заряженные атомы и молекулы.
На основе идеи электролитической диссоциации были даны первые научные определения понятием «кислота» и «основание», согласно которым кислота (например, HCl) это водородосодержащее соединение при диссоциации которого образуются ионы водорода, а основание – например, NaOH, соединение при диссоциации которого образуются ионы гидроксида.
Причины, приводящие к явлению диссоциации, в теории Аррениуса не рассматривались. Не обсуждался также вопрос о том, почему заряженные частицы, на которые должны были бы распространяться законы электростатики, не взаимодействуют друг с другом в растворе [40].
В 1903 году Сванте Аррениус за теорию электролитической диссоциации получил Нобелевскую премию в области химии.
Обычно в книгах по неврологии идёт отсылка к теории Аррениуса, а дальше описываются мембранные потенциалы, как само собой разумеющееся. Но для любопытного читателя замечу, что в теории электролитической диссоциации рассматриваются чисто электрохимические процессы в электролите. Она больше подходит к разборкам в споре между Луиджи Гальвани и Александром Вольта.
В 1890 году Вильгельм Оствальд (1853—1932), продолжая исследования полупроницаемых искусственных плёнок, совместил све́дения об осмосе с положениями теории диссоциации. Он обнаружил, что полупроницаемость плёнок может вызвать не только осмос, но и стать причиной электрических явлений.
Осмос возникает тогда, когда сквозь мембрану приникают относительно мелкие молекулы растворителя (например, воды), но не проходят крупные молекулы растворенного в ней вещества. Но ведь в электролите и ионы могут иметь разные размеры!
Если взять жидкость в сосуде разделить её полупроницаемой плёнкой на две части, в левую и правую части сосуда добавить электролит разной концентрации, и если сквозь плёнку могут проникать только относительно мелкие ионы, например, отрицательные, то после диффузии электролита между левой и правой половинами сосуда возникнет разность электрических потенциалов.
Оствальд также предположил, что свойствами полупроницаемой мембраны можно объяснить возникновение электрических потенциалов мышц, нервов, а также электрических органов рыб. Идея Оствальда, как ни странно, оказалась незамеченной ни биологами, ни физиологами того времени. И только Юлиус Бернштейн (J. Bernstein) спустя десять лет смог по достоинству её оценить.
Теория Чаговца
Но в это десятилетие произошло одно интересное событие. Российский физиолог Василий Юрьевич Чаговец (1897—1941) в 1898 году опубликовал в «Неврологическом вестнике» первую в истории теорию происхождения биоэлектропотенциалов. Чаговец одним из первых применил для объяснения эффекта электрогенеза теорию электролитической диссоциации Аррениуса, поэтому и работу свою назвал: «О применении теории диссоциации растворов электролитов Аррениуса к электрофизиологии».
Опирался он и на представления Лудимара Германа о том, что ток покоя является альтерационным1 током.
Чаговец рассуждал так: если мышцу возбудить, то обмен веществ в участке возбуждения резко возрастёт (по данным Германа, мышца в состоянии те́тануса выделяет в 6,5 раз больше СО2, чем покоящаяся мышца). Следовательно, в этом участке значительно увеличится количество метаболитов, в том числе и угольной кислоты, которая диссоциирует на ионы водорода и ионы СО2.
Эти ионы по закону диффузии потекут от возбуждённого или повреждённого участка к покоящемуся (неповреждённому).
Но подвижность ионов водорода намного выше, чем скорость перемещения ионов СО2, поэтому очень скоро неповреждённый участок приобретает положительный потенциал, повреждённый же – отрицательный.
Теоретический расчёт Чаговца показал, что разность потенциалов между возбуждённым и покоящимся участками должна равняться 0,038 В (при условии, что интенсивности метаболизма в возбуждённой мышце в 6,5 раз выше, чем в покоящейся).
Чаговец проверяет соответствие теоретически предсказанных значений биопотенциалов экспериментальным. Практические измерения дали в среднем величину 0,043 В (при разбросе от 35 до 50 мВ). То есть, величи́ны, вычисленные теоретически и найденные практически, оказались довольно близкими. Расхождения Чаговец объяснил тем, что поперечный разрез является более сильным раздражителем, чем обычное.
Для своего времени теория Чаговца, как видно из этих рассуждений, была хорошо аргументированной. Однако, она содержала ряд спорных моментов. Например, не вполне корректным было число 6.5, указанное Германом. Также, трудно было согласиться с тем, что в электрогенезе играет роль только углекислота, ведь если повреждённую поверхность нейтрализовать щелочным раствором, она тем не менее не меняет своего отрицательного заряда.
К этому следует прибавить, что не были учтены последние открытия Пфеффера в области полупроницаемых мембран (1877).
1 Альтера́ция (от лат. alterare – изменять) – общее название изменения структуры клеток, тканей и органов, сопровождающееся нарушением их жизнедеятельности.
Мембранная гипотеза Бернштейна
В соответствии с теорией Аррениуса, опираясь на идеи Оствальда и исследования явлений осмоса, Бернштейн предположил, что возникновение и проведение нервного импульса обусловлено перемещением ионов между нервным волокном и внеклеточной средой.
Именно Бернштейн, работая тогда вместе с Германом, доказал прямыми экспериментами, что возбуждённый участок поверхности мышцы или нерва на очень короткий промежуток времени приобретает потенциал, отрицательный по отношению к невозбуждённой или неповреждённой поверхности.
В 1902 году Юлиус Бернштейн (Bernstein, 1839—1917) выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К+, и они накапливаются в цитоплазме создавая на поверхности электрический потенциал – потенциал покоя. Согласно этой гипотезы, при возбуждении клетки, её мембрана «повреждается», и ионы К+ выходят из неё до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, а потенциал возвращается к уровню потенциала покоя.
Что такое потенциал покоя, и что такое электрический потенциал? Проще всего это можно пояснить на примере обыкновенной пальчиковой батарейки. У неё есть два контакта (полюса) «плюс» и «минус». Напряжение батарейки 1,5 вольта. И разность потенциалов между плюсом и минусом как раз и составляет полтора вольта. То есть, можно представить, что на одном полюсе батарейки потенциал плюс 0,75 В, а на другом – минус 0,75 В. Разница между плюсом и минусом составит те самые 1,5 В. Разность потенциалов и величина одного потенциала измеряются в вольтах (В).
Напряжение – это разность потенциалов между двумя точками. Напряжение определяется исключительно относительно некоторого уровня, обозначается буквой U.
Потенциал покоя – это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. С точки зрения физика, приведённое выше определение неправильное. Потенциал не может быть разностью потенциалов. Но у медиков принята именно такая терминология. Его средняя величина для нейрона составляет 70 мВ (70*10—3 В).
Отметим, что заряд мембраны измеряется изнутри клетки, а не снаружи. Проще говоря, снаружи вокруг клетки будут преобладать «плюсики», т. е. положительно заряженные ионы, а внутри – «минусики», т. е. отрицательно заряженные.
Согласно идее Бернштейна, в клетке всегда есть электричество, её внутренняя часть заряжена отрицательно по отношению к наружной среде, а эта разность потенциалов и есть причина электрического тока. Поэтому он предположил, что при раздражении в клеточной мембране действительно возникает «дырка», но не реальная, как при разрезе или проколе, а «электрическая», дырка для токов, т. е. мембрана становится проницаемой не только для калия, стремящегося внутрь, но и для других ионов.
Из этой гипотезы напрашивался вывод, определяющий содержание контрольного эксперимента: проверить сопротивление в области возбуждения мембраны, оно должно снижаться за счёт этой «дырки». Бернштейн сделал попытку проверить это предсказание экспериментально. Но продолжить развитие и обоснование своей гипотезы, увы, не успел: его книга с описанием открытия вышла в 1912-м, этот год и считается годом рождения мембранной теории биопотенциалов, вскоре началась Первая мировая война, а в 1917 году Бернштейн умер.
Гипотеза Бернштейна о генерации потенциала покоя на клеточной мембране была встречена в научном сообществе без особого энтузиазма. В глазах современников она выглядела не более чем оригинальной гипотезой и требовала экспериментальных подтверждений. У скептиков главными аргументами были: во-первых, отсутствие экспериментальных доказательств существования само́й мембраны – её увидели в электронный микроскоп лишь в 1950 году, и во-вторых, наличие ионов калия внутри клетки подтверждалось лишь косвенными данными.
Из трёх основных «действующих лиц» мембранной теории Бернштейна: мембраны, наружной среды и внутриклеточной среды, достаточно хорошо на тот момент была исследована лишь наружная среда, и не только потому, что она была наиболее доступной. [14]
Химическим составом среды, окружающей клетки организма, биологам уже давно приходилось заниматься. При проведении экспериментов на изолированных о́рганах их следует хранить в специальном растворе. Например, лягушачью лапку нельзя подолгу оставлять просто на воздухе – она высохнет и перестанет работать, но нельзя и поместить её в чистую воду – под действием осмоса клетки препарата погибнут.
Таким образом, важный для мембранной теории солевой – а значит и ионный – состав внеклеточных жидкостей был хорошо известен врачам. Оказалось, что основу этой жидкости составляет 9% раствор простой поваренной соли NaCl (физраствор).
Примечателен и очень важен для мембранной теории тот факт, что, соотношение концентраций ионов натрия и калия в среде, окружающей клетки организма, примерно одинаково для всех животных – от медузы до человека. Независимо от концентрации, количество калия в растворе примерно в 50 раз меньше, чем натрия. Так что, у всех животных межклеточная среда по существу представляет собой в бо́льшей или меньшей мере разбавленную морскую воду.
Между тем, в 1908 году была опубликована модель биоэлектрогенеза Вальтера Нернста (Nernst, 1864—1941). Биоэлектрогенез – это процесс генерации электричества живыми организмами.
Нернст взял сосуд с растворами КCl разной концентрации, разделёнными полупроницаемой мембраной. Из-за различия в проницаемости мембраны для катионов К+ и анионов Сl-, за определённое время через мембрану проходит гораздо больше ионов калия, чем хлора. В результате, в растворе с низкой концентрацией возникнет избыток К+, и раствор приобретёт положительный заряд, а в растворе с более высокой концентрацией остаётся больше Cl-, и этот раствор станет отрицательно заряженным. Так как эти заряды притягивают друг друга, то на мембране возникнет двойной электрический слой – по одну сторону скопятся положительные заряды (ионы К), а по другую – отрицательные (ионы Cl). Вследствие этого на мембране возникнет разность потенциалов. Этот постоянный потенциал назвали диффузионным. Справедливости ради заметим, что ещё в 1890 году Вильгельм Оствальд провёл аналогичный опыт. Но важной заслугой Нернста стало математическое описание этого процесса (формула Нернста):
η = μ+zψF
где
μ – химический потенциал,
z – валентность вещества,
ψ – электрический потенциал фазы,
F – число Фарадея.
Нернст изучал поведение электролитов при пропускании электрического тока и открыл закон, устанавливающий зависимость между разностью потенциалов и ионной концентрацией. Уравнение Нернста позволяет рассчитать максимальный рабочий потенциал, который может быть получен в результате электрохимического взаимодействия, при заданных давлении и температуре. Таким образом, этот закон связывает термодинамику с электрохимической теорией в области решения проблем, касающихся сильно разбавленных растворов.
Именно по этой формуле Бернштейн в 1912 году рассчитал величину потенциала покоя для К+ совпавшую с экспериментально измеренным потенциалом между саркоплазмой мышцы и окружающей средой, который составлял около – 70 мВ.
Оставалось экспериментально доказать наличие биоэлектрогенеза в живой клетке.
До современного представления о распространении нервного импульса оставались считаные шаги, но ещё многие годы исследования.
«Язык головного мозга». Гассер и Эрлангер
Ещё в 1868 году молодой немецкий физиолог Юлиус Бернштейн с помощью изобретённого им дифференциального реотома сумел определить форму нервного импульса. Она оказалась колоколообразной.
Спустя несколько десятилетий, в начале 1900-х американский учёный Герберт Гассер (Gasser Herbert Spencer, 1888—1963) вместе с коллегой Жозефом Эрлангером (Joseph Erlanger, 1874 – 1965) задались целью усиления и визуализации электрических сигналов отдельных нервных волокон.
Гассер понимал, что для регистрации амплитуды нервного импульса нужен более современный прибор, чем гальванометр. Этот прибор должен был одновременно прочитать все параметры электрического сигнала, визуализировать и записать их на ленту. Говоря современным языком, учёный нуждался в осциллографе.
Различные варианты осциллографов начали появляться с 1880 года и к 1920 году прибор представлял собой катодную трубку – аналог электроннолучевых кинескопов, которые применялись в наших телевизорах до появления плазм и ЖК-экранов.
У Гассера не получилось договориться с компанией-производителем и получить их прибор, поэтому они с Эрлангером создали собственную электровакуумную трубку из колбы для дистилляции воды. Именно таким самодельным осциллографом учёные зарегистрировали первую в мире осциллограмму с записью электрических импульсов, возникающих в нервных клетках.
В периферической нервной системе отдельные волокна объединены в нервные стволы (нервы). В одном нерве могут быть тысячи нервных волокон. Волокна в нервах могут быть миелиновыми и безмиелиновыми. В естественных условиях каждое волокно возбуждается от своего источника, и электрические потенциалы в них проводятся несогласованно. Кроме того, по чувствительным (афферентным) и двигательным (эфферентным) волокнам импульсы бегут на встречу друг другу. Результирующая электрическая активность нерва создаётся электрической активностью всех составляющих его волокон. В связи с этим анализ суммарной электрической активности нерва (нейрограммы) представлял трудную задачу. Учёные поначалу зафиксировали только «белый шум» на экране осциллографа, но догадались, что – это не что иное, как совокупность электрических импульсов от множества нейронов. Ведь измерения проводились не на отдельном нейроне, а на нерве, похожем на многожильный кабель.
Они предположили, а потом и доказали, что скорость проведения электрического потенциала зависит от толщины нервного волокна. Чем тот толще, тем быстрее способен передавать сигнал. Такое предположение впервые выдвинул шведский физиолог Густав Гётлин ещё в 1907 году, но с тех пор никто не пытался проверить или опровергнуть его.
Для наглядности классификации Гассер свёл все параметры в единую таблицу, которую и поныне можно найти в медицинских справочниках.
Эти опыты значительно продвинули учёных в понимании механизма прохождения нервного импульса и легли в основу нейрофизиологии. Все полученные све́дения позднее были применены в модели нервной проводимости, разработанной Аланом Ходжкином и Эндрю Хаксли в 1952 году.
В 1937 годe Гассер и Эрлангер опубликовали совместную книгу «Электрическая регистрация нервной деятельности». А в 1944 им вручили Нобелевскую премию «за открытия, имеющие отношение к высокодифференцированным функциям отдельных нервных волокон». Церемонию награждения во время Второй мировой войны не проводили, только по радио транслировали поздравительную речь. Но в 1947 году они всё же прочитали свои Нобелевские лекции «Нервные волокна млекопитающих» в Стокгольме.
Кабельная теория и подводные кабельные линии
Проводя своё расследование источников современных представлений о нейроне, я раскопал и эту историю. Она показалась мне настолько интересной, что рискнул вам о ней рассказать.
О строительстве первых трансатлантических телеграфных линий написал тогда ещё не фантаст, но уже замечательный писатель Артур Кларк в своей научно-популярной книге «Голос через океан» [41] в 1940-х годах. Книга настолько интересна, что вероятно повлияла и на исследователей нервного импульса. Тем более что содержала едва ли не прямое на то указание, например, «…этот кабель – жизненный нерв современного общества, подобный нервам живого организма, это неотъемлемая часть общей мировой системы связи…» или «Человеческое общество совершенствует свою „нервную систему“, стараясь „чувствовать“ каждую часть своего организма… Связь и есть та нервная система, которая позволяет знать, что делается в данный момент в любой части мира.»
Стоит отметить, что в книге А. Кларка описаны события более чем полутора вековой давности. Как когда-то древние римляне сравнивали нервы с виадуками, так и современники описываемых событий сравнивали нервные импульсы с последними технологическими достижениями. Поэтому не удивительно, что компетентнейший учёный тех лет Герман Гельмгольц писал: «Нервные волокна часто сравнивают с телеграфными проводами, пересекающими местность, и это сравнение хорошо приспособлено для иллюстрации удивительных и важных особенностей их образа действия. В телеграфной сети везде мы обнаруживаем те же медные или стальные провода, несущие только один вид движения, поток электричества, но вызывающие самые разные результаты на разных станциях в соответствии с дополнительной аппаратурой, с которой провода соединены. На одной станции эффект состоит в звонке колокольчика, на другой сигнал просто передаётся дальше, на третьей вступает в работу записывающий аппарат. …Говоря коротко, каждое из… различных действий, вызываемых электричеством, может быть вызвано и передано проводом в любую необходимую точку. При этом в проводе происходит один и тот же процесс, приводящий к самым разным последствиям. …Та разница, которую мы видим при возбуждении различных нервов, заключается только в разнице самих органов, к которым присоединён нерв и которым передаётся состояние возбуждения». [42]
Поэтому, возможно, в 1940-х годах, с подачи Лудимара Германа, Алан Ходжкин со своими коллегами математически применили кабельную теорию Томсона (Кельвина) к аксонам беспозвоночных, тем самым положив начало современному математическому описанию и моделированию нейронов.
С чего всё началось. Первый подводный кабель, передающий электрический сигнал, был проложен в Мюнхене по дну реки Изар в 1811-м году. Идея принадлежала немецкому врачу и изобретателю Зёммерингу (Samuel T. von Soemmering).
Однако из-за отсутствия технологии эффективной гидроизоляции долгое использование подобного кабеля не представлялось возможным. Лишь изобретение в 1847 году Сименсом технологии изготовления изоляции из гуттаперчи1 позволило начать работы по прокладке кабеля между Кале и Дувром (который, кстати, разорвался после пересылки первой же телеграммы, а год спустя после замены его новым армированным кабелем, тоже прослужил недолго).
В 1856 году было основано акционерное общество «Atlantic Telegraph Company», которое в 1857 году приступило к укладке 4500-километрового армированного телеграфного кабеля через Атлантический океан. Кабель, весивший около 550 кг/км, состоял из семи медных проводов, покрытых тремя слоями гуттаперчи и защитной оболочкой из стальных тросов.
В августе 1857 года корабли «Агамемнон» и «Ниагара» начали прокладку от юго-западного берега Ирландии, однако, из-за разрыва кабеля работу пришлось отложить на год.
Вторая попытка была предпринята летом 1858 года. На этот раз решили начать прокладку от точки стыковки в океане, примерно посередине между Ирландией и Ньюфаундлендом. 26 июля «Агамемнон» и «Ниагара», каждый со своей половиной кабеля на борту, встретившись в Атлантическом океане, соединили половины кабеля, опустили его в воду и начали укладку в разных направлениях.
В процессе прокладки кабель несколько раз разрывался, и кораблям приходилось возвращаться, чтобы начать заново.
5 августа корабли достигли своих пунктов назначения – островов Валентия и Ньюфаундленд, – и была установлена первая трансатлантическая телеграфная линия, соединяющая Старый и Новый Свет. 16 августа 1858 года королева Великобритании Виктория и президент США Джеймс Бьюкенен обменялись поздравительными телеграммами. Приветствие английской королевы состояло из 103 слов, передача которых длилась 16 часов! Никто до этого и предположить не мог, что скорость передачи будет столь далека от скорости света. Телеграфировать в таком медленном темпе приходилось потому, что из-за огромной электрической ёмкости и сопротивления длинного кабеля короткие импульсы тока «расплывались» на приёмном конце подобно чернильным кляксам на промокашке.
Уже в сентябре 1858 года связь была нарушена. Видимо, ввиду недостаточной гидроизоляции, кабель был испорчен коррозией. Другой возможной причиной разрушения стали слишком высокие напряжения – 2000 вольт, подаваемые на линию с английской стороны с целью ускорения передачи.
В пятидесятых годах XIX века Уильям Томсон (Кельвин, William Thomson, 1-st Baron Kelvin, 1824 – 1907) заинтересовался проблемами трансатлантической телеграфии. Вообще-то, предметом его исследований были процессы, происходящие в проводнике между моментом подачи на него напряжения и тем моментом, когда это напряжение достигнет заданной величины. Побуждаемый собственным любопытством и неудачами первых пионеров-практиков, Томсон теоретически исследовал вопрос распространения электрических импульсов по кабелю и пришёл к заключениям величайшей практической важности, давшим впоследствии возможность осуществить телеграфирование через океан.
Многие ошибочно полагают, будто электрический ток идёт по проводу со скоростью света, равной 300 000 километров в секунду. На самом же деле ток течёт по проводам в несколько раз медленнее, чем распространяется свет.
Скорость тока в кабеле тем меньше, чем больше его электрическая ёмкость, измеряемая в Фарадах. К счастью для телеграфной связи, на первых порах это явление не оказывало практически никакого влияния. Ёмкость коротких линий была столь мала, что сигналы проходили по ним без сколько-нибудь заметной задержки. Но когда были проложены трансконтинентальные кабели, эта задержка послужила источником многих проблем.
Исследования Томсона привели к открытию его знаменитого «закона квадратов», согласно которому скорость телеграфирования по кабелю обратно пропорциональна квадрату его длины. Иначе говоря, увеличив длину кабеля, например, в 10 раз, мы получим снижение скорости передачи в 100 раз. Безусловно, что такое открытие имело исключительно важное значение для подводного телеграфирования на дальние расстояния.
Компенсировать уменьшение скорости передачи по длинным телеграфным линиям инженеры того времени могли исключительно, увеличивая диаметр токопроводящей жилы и улучшая гидроизоляцию.
Спустя шесть лет, в 1864 году началась укладка 5100-километрового кабеля с улучшенной изоляцией, 7-проволочная медная жила была изолирована четырьмя слоями гуттаперчи и покрыта сначала пропитанной пенькой, затем десятью стальными бронепроволоками, каждая из которых предварительно обмотана слоем пеньки. На береговые концы трансатлантического кабеля конструкции 1865 года поверх глубоководного кабеля наложены пеньковая подушка и усиленная броня, состоящая из 12 тросов, каждый из которых скручен из трёх стальных проволок [41].
1 Гуттаперча – смола, добываемая из растения pertja, высокомолекулярный углеводород, идентичный по химическому составу с натуральным каучуком

Рисунок 32. Трансатлантический телеграфный кабель конструкции 1865—1866 гг. Разделанный конец и поперечное сечение кабеля.
В качестве кабелеукладчика было решено задействовать крупнейшее судно тех времён – британский пароход «Грейт Истерн» водоизмещением 32 тысячи тонн. 31 июля 1865 года при укладке опять произошёл обрыв кабеля. Лишь в 1866 году со второй попытки удалось уложить кабель, который обеспечил долговременную телеграфную связь между Европой и Америкой.
Спустя 10 лет с помощью значительно лучше изолированного кабеля удалось проложить сразу несколько трансатлантических телеграфных линий с большей долговечностью. И к 1919 году число кабелей достигло 13, большинство из них принадлежали Великобритании.
А как же скорость и качество передачи данных? Предложенное Томсоном решение улучшило качество связи, но, увы, оно ещё долгие годы оставалось крайне низким.
В чём же заслуги Томсона? Во-первых, он объяснил причину возникающих проблем. Всё дело в том, что морская вода, несмотря на все усилия конструкторов кабеля попадала под его внешнюю броню и это приводило к значительному увеличению электрической ёмкости между ней и центральной жилой. При этом внешняя морская среда становилась участником процесса передачи сигнала, чего не происходило на суше. Во-вторых, Томсон предложил оригинальную идею, которая не решила возникших проблем, зато позволила их обойти. Суть её в том, чтобы приёмное устройство фиксировало не весь сигнал: точка или тире, а только его начальный фронт. Для этого нужно было вместо увеличения мощности отправляемого сигнала, усилить чувствительность принимающего устройства. Эта задача была частично решена благодаря внедрению сконструированного Томсоном же чувствительного зеркального гальванометра.
Окончательно все проблемы были сняты только при строительстве трансатлантического телефонного кабеля TAT-1, который был проложен почти сто лет спустя между городами Обаном (Шотландия) и Кларенвиллем (Ньюфаундленд) в 1956 г. Эта телефонная линия содержала 51 усилитель, расположенный на расстоянии 70 км друг от друга. Именно внедрение промежуточных усилителей-ретрансляторов позволило осуществлять телеграфное, и даже телефонное, сообщение приемлемого качества. До того момента, начиная с 1927 года, телефонное сообщение между Старым и Новым Светом осуществлялось по радио в длинноволновом диапазоне.
Вот такова предыстория кабельной теории распространения нервного импульса.
Кабельная теория нервного импульса
Впоследствии распространилось мнение, что подводный кабель схож по своим свойствам с нервным волокном. Он имеет токопроводящие сердцевины, покрытые изолирующей оболочкой, и окружён морской водой. Так же и нейроны находятся в соляном растворе. И, поскольку изоляция кабеля не является совершенной, то существует конечное сопротивление утечки через изолятор. Видимо, учёные увидели в этом аналогию с проницаемостью мембраны. Значительные количественные и качественные различия между кабелем и нейроном состоящие в том, что кабельные жилы сделаны из меди и являются намного лучшим проводником, чем электролитический раствор нейрона, а также то, что кабельное покрытие – это настоящий изолятор, в отличие от мембраны клетки, не были приняты во внимание.
Тот факт, что по центральной жиле кабеля течёт ток, а в нейроне мембранный потенциал распространяется по мембране, также не учли. Эту основу основ электрических свойств нейрона попросту проигнорировали.
Основная задача кабельной теории Томсона – объяснить причины замедления сигнала и увеличить дальность передачи телеграфных сообщений в очень длинных кабельных системах, тоже была отодвинута в сторону. Как и несопоставимость масштабов процессов, происходящих в тысячекилометровых кабельных линиях и сравнительно коротких нейронах. Зато стали активно исследовать процесс затухания нервного импульса в аксоне, в то время как один из основополагающих законов распространения нервного импульса звучит так: Всё или ничего. То есть потенциал действия не может затухать, он либо есть, либо его нет. Но, никакой другой, более подходящей теории на тот момент не нашлось.
Теория местных токов
Что такое местные токи, на основе которых строится кабельная теория распространения потенциала действия? Давайте попробуем разобраться.
Итак, предположим, к аксону приложен стимулирующий электрод. В результате в точке его приложения возникнет некоторый электрический потенциал. Но как этот потенциал будет изменяться по мере удаления от точки раздражения? Ответ найден экспериментально – он уменьшается. Дальше логика исследователей такова: поскольку потенциалы, измеренные в двух разонудаленных точках, отличаются, то существует некоторая разность потенциалов между ними. А дальше совсем просто. Зная разность потенциалов – U и измерив сопротивление мембраны R, по закону Ома вычисляем ток I=U/R.
Чудо! Скажет кто-то. Но это «чудо» объясняют электрикам 1-го (низшего) разряда на инструктаже по технике безопасности. «Подходить к находящемуся под напряжением проводу, лежащему на земле, следует мелкими шажками». Явление это так и называется «шаговое электричество».
Вот эти-то токи, возникающие вблизи возбуждённой области, Лудимар Герман назвал «местными токами», поэтому и его теорию называют теорией местных токов. [23]
Идея Германа состояла в том, что токи, возникающие на некотором отрезке возбуждённого волокна, выступают в роли раздражителя для соседних точек того же са́мого волокна; в результате возбуждение переходит на соседнюю область, которая, в свою очередь, становится раздражителем для следующего ещё невозбуждённого участка волокна, и т. д. И точка максимального потенциала, о которой мы говорили в самом начале начинает двигаться по мембране. «Ну это вряд ли», – скажет вам электрик 1-го разряда. Напряжение будет уменьшаться и уменьшаться по мере удаления, пока совсем не сойдёт на нет, а вместе с ним и ток.

Рисунок 33. Измерение местных токов.
Было установлено, что значение потенциала уменьшается по мере удаления от источника возбуждения по экспоненциальному закону:
ψ i= ψ 0 * e -1/λ
где ψ0 – значение потенциала в точке возбуждения, ψl – значение потенциала в точке, расположенной на расстоянии λ от источника возбуждения, λ – константа длины нервного волокна, равная расстоянию, на котором величина потенциала убывает в e раз (е=2,718281828…).
Константа λ зависит от удельного электрического сопротивления оболочки нервного волокна ρm, удельного электрического сопротивления цитоплазмы ρi и радиуса нервного волокна r:
Чем больше λ, тем больше скорость распространения нервного возбуждения. Как следует из приведённой выше формулы, λ тем больше, чем больше радиус нервного волокна и чем больше удельное электрическое сопротивление мембраны нервного волокна.
Вот тут-то и пригодилась кабельная теория Томсона с её волновыми процессами. Чтобы они возникли надо было добавить в схему нелинейные электрические элементы – ёмкость (С) и индуктивность (L). С индуктивностью вышла промашка, в трансатлантическом кабеле она была, а вот в нейроне, увы, нет (слишком короток). Зато с ёмкостью всё сложилось складно. Именно ёмкость между внутренней жилой и внешней средой стала причиной рождения кабельной теории Томсона. У клетки также есть внутренняя среда и внешнее межклеточное пространство разделённое диэлектриком-мембраной. Стоп, кажется, раньше мембрана не была диэлектриком, а вполне успешно обладала вполне конкретным сопротивлением R. Ну ничего страшного, решили учёные. Значит, мембрана обладает двумя сопротивлениями: одно вдоль, а другое поперёк мембраны и назвали его Rm.
Стоит отметить, в 1923 году (голландские физиологи) И. Гортер и А. Грендел сделали предположение, что липиды в мембране располагаются в два слоя.
Они провели оригинальный эксперимент: поместили эритроциты в пресную воду, при этом по градиенту концентрации вода должна проходить через мембрану в клетку. При этом эритроциты разбухают, мембраны их лопаются и клетки теряют содержимое. Остаются лишь прозрачные наружные мембраны. Это позволило прояснить структуру клеточных мембран. Липиды были экстрагированы ацетоном из мембран эритроцитов. Помещённые на поверхность воды липиды образовывали пятна толщиной в одну молекулу. Измерив площадь этого слоя, авторы определили, что она вдвое больше, чем площадь всех мембран эритроцитов, из которых липиды были извлечены. Было высказано предположение, что липиды в мембранах располагаются в два слоя.
А двойной фосфолипидный слой биологической мембраны вскоре был уподоблен конденсатору, в котором слои́ играют роль обкладок. Это предположение сделали биологи К.С.Кол и Г. Кертис в своих исследованиях электрических параметров биологических мембран в 1939 году. Именно они вычислили высокое электрическое сопротивление монослоя липидов Rm = 107 Ом/м2 и большую электрическую ёмкость бислоя мембраны С = 10—2 Ф/м2. Кстати, эти два талантливых учёных вычислили электрическую ёмкость клетки вцелом, а эквивалентную схему с множеством RC-контуров предложили Ходжкин и Хаксли.
Оставалась последняя неувязка – отсутствие в цепи индуктивности. Без неё не работает электрический колебательный контур. Ну и ладно, пусть роль индуктивности играет то самое поперечное сопротивление Rm.
Так возникла первая эквивалентная электрическая схема нервного волокна.

Рисунок 34. Эквивалентный контур, гипотетического кабеля. Сопротивления и ёмкости соединены через продольные наружные и внутренние сопротивления. Мембранное сопротивление RM, продольное сопротивление Rn и ёмкость СM произвольно расчленены на отдельные элементы 0, 1, 2, 3, 4. Цветные стрелки показывают направление тока.
Ныне теория местных токов Германа общепризнана и составляет основу электробиологии наряду с мембранной теорией Бернштейна.
И от этой точки начали расти и множиться теории и объяснения процессов, происходящих при распространении нервного импульса.
Безусловно, наука только тогда становится наукой, когда исследуемые процессы описаны на языке математики.
Но в результате спорная, в общем-то, гипотеза получила такое математическое углубление, что уже просто не может не вызывать абсолютного к себе доверия.
Интересно, если бы Кларк написал свою книгу о строительстве нефте– и газопроводов, история пошла бы другим путём? И мы сравнивали бы сейчас нервные волокна с водопроводными трубами?
Но, как бы то ни было, именно предложенная в 1905 году Германом кабельная теория, дала первое объяснение механизма передачи возбуждении по нервному волокну.
Ходжкин и Хаксли (Hodgkin & Huxley)
Ходжкин и Хаксли получили Нобелевскую премию по физиологии и медицине 1963 года «за открытия, касающиеся ионных механизмов, участвующих в возбуждении и торможении в периферическом и центральном участках мембраны нервной клетки».
Но что конкретно они сделали, в чём их заслуга? Условно, в работах Ходжкина и Хаксли можно выделить три основные направления:
Они развили и дополнили мембранную теорию Бернштейна объяснив, как распространяется вдоль нервного волокна бегущая волна ПД1.
Объяснили, как восстанавливается ионный баланс, обеспечивающий ПП, после прохождения ПД.
Создали математическую модель описывающую генерацию и распространение потенциалов действия в нейронах.
1 ПД – потенциал действия, ПП – потенциал покоя
Развитие мембранной теории
Предсказание Бернштейна об изменении проницаемости мембраны при возбуждении удалось проверить только спустя четверть века, в 1938 году. Хотя возобновившееся после Первой мировой войны изучение свойств мембраны совершенствовалось, технические трудности были слишком велики: ведь надо было уловить изменение электрического потенциала, которое длится всего несколько миллисекунд!
Первого прорыва достигли в 1937 году американские биологи К.С.Кол и Г. Кертис, которые догадались вместо традиционных нерва или мышцы лягушки исследовать клетку водоросли. То, что растения способны к генерации электрических потенциалов, учёным известно было давно. Знали они и то, что все электрические процессы у растений идут значительно медленнее. Оставалось найти только клетку побольше – такая, с диаметром около полмиллиметра, нашлась у водоросли нителлы.
На этом неожиданном объекте и провели свой опыт Кол и Кертис. Они обнаружили, что при возбуждении сопротивление мембраны, как и предсказывалось, уменьшается, правда, не до нуля, но существенно, почти в 200 раз.
Через год, в 1938 году, одновременно две группы исследователей в двух местах – К.С.Кол и Г. Кертис, работавшие на морской станции Океанологического института в Вудс-Холе, и Ходжкин с Хаксли, работавшие на морской станции в Плимуте, измерили потенциал действия и потенциал покоя на новом модельном объекте – гигантском аксоне кальмара.
Гигантские аксоны есть у многих беспозвоночных. Но у кальмара он особенно крупный – диаметр его у обычных кальмаров рода Loligo (с длиной тела 30—40 см) достигает 1 мм, а у некоторых видов ещё в 2—3 раза больше.
Гигантские аксоны были описаны ещё в начале ХХ века; но об открытии забыли, и все считали их кровеносными сосудами. «Переоткрыл» гигантские аксоны зоолог Джон Янг в 1936 году. И не только переоткрыл, но и предложил физиологам использовать их для работы. Совет оказался очень удачным. Наилучшим образом им воспользовались Алан Ходжкин и Эндрю Хаксли, получившие за свою работу Нобелевскую премию.
На этом этапе Алану Ходжкину и Эндрю Хаксли по образцу экспериментов Кеннета Кола и Говарда Кертиса удаётся создать миниатюрные электроды и ввести их в аксон. Обе группы с помощью внутриклеточных электродов определяют полный потенциал покоя на гигантских нервных волокнах кальмара и публикуют свои результаты в 1939 и 1940 годах.
Как в своё время Первая мировая война помешала Бернштейну исследовать гипотезу «дырок», так начавшаяся 1 сентября 1939 года Вторая мировая вынудила Ходжкина и Хаксли отложить свои исследования. Во время войны Ходжкин участвовал в разработке радиолокационных станций.
Двухэлектродная фиксация потенциала
Только по окончании войны, в конце 40-х – начале 50-х годов, натриевая гипотеза приобретает современные черты, формируется натриевая теория.
Ведущую роль в дальнейших исследованиях сыграл новый метод фиксации потенциала с помощью двух электродов (в англоязычной литературе TEVC – two-electrode voltage clamp), разработанный в конце 1930-х годов К. С. Колом и Дж. Мармонтом. Этот метод позволял измерять не только потенциал, но и токи при заданном значении мембранного потенциала.
Идея метода «зажима напряжения» приписывается Кеннету Колу и Джорджу Мармонту. Весной 1947 года Кол обнаружил, что можно использовать два электрода и цепь обратной связи для поддержания мембранного потенциала клетки на заданном экспериментатором уровне.
Два электрода были сделаны из тонких проводов, скрученных вокруг изоляционного стержня. Поскольку подобные электроды можно было вставить только в самые большие клетки, первые электрофизиологические эксперименты проводились почти исключительно на аксонах кальмаров.
В клетку помещали два электрода, один из которых измерял потенциал относительно внеклеточного электрода сравнения и передавал его значение на специальный усилитель, который сравнивал измеренный потенциал со значением командного. Это устройство вычисляло величину тока, необходимого для компенсации этой разницы потенциалов, и подавало через второй внутриклеточный электрод ток такой величины, чтобы потенциал на мембране клетки стал равен Vcmd (Vm = Vcmd). По амплитуде тока, необходимого для компенсации сдвига потенциала до Vcmd, можно было судить о токе через мембрану при данном значении мембранного потенциала. Ток при данном значении потенциала равен току, подаваемому на второй электрод, взятому с обратным знаком.

Рисунок 35. Схема фиксации потенциала с помощью двух электродов
Гигантский аксон кальмара стал первым препаратом, который мог быть использован для измерения напряжения и трансмембранного тока, что и стало основой пионерских экспериментов Ходжкина и Хаксли со свойствами потенциала действия. Летом 1952 года Алан Ходжкин и Эндрю Хаксли опубликовали пять статей, описывающих, как ионные токи вызывают потенциал действия.
PS Метод двухэлектродной записи пережил множество усовершенствований и используется по сей день. А внедрение стеклянных хлорсеребряных электродов, заполненных солевым раствором, позволило использовать метод на более мелких объектах.
Режим фиксации разницы потенциалов на мембране (voltage clamp) идеально подходит для изучения электрофизиологических свойств ионных каналов мембраны. Но иногда требуется решение обратной задачи – записи потенциала при фиксированном значении тока, поэтому большинство современных устройств позволяет работать ещё и в режиме фиксации тока – current clamp. Во многом эти два режима зеркальны: в current clamp на постоянном уровне поддерживается ток, а потенциал записывается, а в voltage clamp – наоборот.
Овершут
Дальнейшие исследования показали, что вариант с «дыркой» в мембране предложенный Ю. Бернштейном не состоятелен: возникновение потенциала действия невозможно объяснить простым «закорачиванием» мембраны. Более того, эксперименты с измерением сопротивления мембраны показали, что в реальности ПД оказался заметно больше ПП.
Надо было как-то объяснить возникновение этого «дополнительного» потенциала. На самом деле для решения этой задачи все данные были уже известны, основные идеи высказаны – оставалось только их сопоставить и сделать решающий вывод.
Суть новой гипотезы заключалась в следующем. Кроме основного потенциала покоя, создаваемого ионами калия внутри клетки, существует ещё один «встречный» потенциал, создаваемый ионами натрия, находящимися во внеклеточной среде. Эти потенциалы суммируются. Пока нейрон пребывает в состоянии покоя, потенциалзависимые каналы мембраны закрыты и поддерживается равновесный суммарный ПП. Когда в результате возбуждения мембранный потенциал увеличивается до порогового уровня, например, с -70 до -55 милливольт, первыми открываются потенциалзависимые натриевые каналы, и ионы натрия устремляются внутрь клетки, вызывая краткое, но резкое увеличение количества положительных зарядов и поднимая мембранный потенциал до +40 милливольт. В ответ на это изменение мембранного потенциала натриевые каналы, открывшись на некоторое время, закрываются, а потенциалзависимые калиевые каналы ненадолго открываются, увеличивая отток положительно заряженных ионов калия из клетки и быстро возвращая мембранный потенциал к состоянию покоя —70 милливольт.
Совсем нелегко было прийти к таким представлениям. И вот почему: диаметр иона натрия в растворе раза в полтора больше диаметра ионов калия и хлора. И совершенно необъяснимо, как бо́льший по размеру ион проходит там, где не может пройти меньший1.
1 О том, как в 1998 разрешена эта загадка узнаем в главе «Исследование биологических мембран»

Рисунок 36. Модель потенциала действия Ходжкина – Хаксли.
Нужно было найти новое объяснение механизма перехода ионов через мембраны. Только осмосом движение ионов через мембрану не объяснить. И тогда была высказана идея, что ионы могут пересекать мембрану с помощью пока неизвестных белковых молекул. Причём эти «молекулы-такси» хорошо различают своих пассажиров, и никогда не путают ионы натрия с ионами калия.
Так элегантно были объяснены значения потенциалов покоя и действия. А следующим шагом стало объяснение распространения нервного импульса путём последовательного открытия натриевого и калиевого каналов мембраны.
Алан Ходжкин прекрасно разбирался в электричестве и дал такую иллюстрацию распространения сигнала в нерве. «Если специалист по электричеству посмотрит на нервную систему, то сразу увидит, что передача сигнала по нервным волокнам является огромной проблемой. Диаметр аксона в нерве варьирует от 0,1 до 20 микрон. Внутреннее содержимое содержит ионы и является неплохим проводником электричества. Однако, волокно невелико и его продольное сопротивление очень большое. Простой расчёт показывает, что в волокне диаметром 1 микрон и сопротивлением 100 Ом/см удельное сопротивление составит около 1010 Ом/см. Это означает, что электрическое сопротивление маленького нервного волокна длиной в 1 метр равно сопротивлению 1010 миль 0,2 мм медной проволоки, то есть проволоки длиной в десять раз больше, чем от Земли до планеты Сатурн» [43].
Требовалось найти другое обоснование электрической природы нервного импульса. И надо сказать, он его нашёл. И, решение это, не менее элегантно, чем объяснение овершута.
Ходжкин и Хаксли совместили теорию местных токов, предложенную Лудимаром Германом, кабельную теорию Томсона и собственную мембранную теорию. Ив вот что у них получилось:
После того как потенциал действия возникает на одном участке аксона, создаваемые при этом ионные токи возбуждают соседние участки, вызывая изменение потенциала действия и на них. Происходящая в результате цепная реакция обеспечивает распространение потенциала действия по всей длине нервного волокна от места, где он был вызван первоначально, до терминалий, предающих сигнал другому нейрону (или мышечной клетке).
Возможно, такая схема и работала бы в идеальных условиях, где нет потерь, так почти бесконечно распространялась бы волна от брошенного в океан камня. В реальных же условиях существуют утечки и другие явления, приводящие к тому, что на каждом следующем участке аксона реполяризация будет уменьшаться и бегущая волна постепенно затухнет. И это не единственная проблема.
Возник ряд сложных и до поры до времени неразрешимых вопросов:
– Почему амплитуда нервного импульса не уменьшается в процессе распространения (НИ не затухает)?
–Как восстанавливается исходный потенциал на мембране после прохождения НИ?
Как проходят ионы натрия и калия через мембрану?
В результате Ходжкин и Хаксли, предложили максимально полное на тот момент биофизическое описание потенциала действия, хотя методы исследования молекулярных механизмов нервного импульса стали доступными только в 80-х годах XX века.
Модели подобные Модели Ходжкина – Хаксли были созданы впоследствии и для других электрохимически возбуждаемых клеток – например, для сердечных миоцитов. И вот что примечательно, во многих русскоязычных источниках, все модели такого рода описывают как автоволновые процессы в активных средах. Более того пример автоволновых процессов в биологии напрямую отсылает читателей к модели Ходжкина – Хаксли.
Для того чтобы понять современное представление о распространении нервного импульса нам придётся сделать отступление и узнать кое-что про автоволновые процессы.
Автоволны
Автоволнами называют волны, распространяющиеся в активных средах, т.е. в средах с распределёнными запасами энергии. Простейшим примером активной среды является бикфордов шнур. Ещё Лудимар Герман предложил на рубеже XIX—XX веков этот пример для вероятного описания нервного импульса. В случае бикфордова шнура запасённая в нём химическая энергия в процессе горения расходуется на розжиг соседних ещё не сгоревших участков шнура. В результате возникает волна горения, распространяющаяся вдоль шнура. Подойдут в качестве примера и падающие костяшки домино, и распространяющийся степной пожар.
Обобщая можно сказать, что автоволны представляют собой самоподдерживающиеся сигналы, которые запускают процессы локального высвобождения запасённой в среде энергии, затрачивающейся на запуск аналогичных процессов в соседних областях.
Мы рассмотрели, пример в котором распространяющийся фронт пламени необратимо переключает её в «сгоревшее» состояние. Особый интерес для исследователей представляют так называемые активные среды с восстановлением, в которых протекают медленные процессы, возвращающие среду из низкоэнергетического состояния (после пробега автоволны) в исходное.
Разберём пример активных сред с восстановлением: горелку с медленно подводящими топливо фитилями. Представим себе горелку, устроенную следующим образом. В листе металла на близком расстоянии друг от друга просверлены отверстия, в которые вставлены полосы асбеста. Концы этих полос погружены в ванну с густым маслом. Асбест не горит, но когда он пропитывается маслом, то представляет собой фитиль, который можно поджечь. Скорость горения асбестового фитиля, пропитанного маслом, выше скорости поступления горючего вещества (масла). Поэтому фитиль через некоторое время погаснет. После этого за счёт диффузии он вновь пропитается маслом, и его вновь можно поджечь и т. д. Таким образом, фитиль может находиться в трёх состояниях: горение; пауза (рефрактерный период), когда засасывается масло; готовность вновь вспыхнуть после поджога (стадия покоя). Если в такой демонстрационной горелке поджечь один из фитилей, то от него загорится соседний. Первый фитиль вскоре погаснет (выгорит масло) но к этому времени по горелке уже побежит фронт пламени. Так технически остроумно была реализована активная среда с восстановлением: каждый её элемент (фитиль) может в отличие от бикфордова шнура вспыхнуть не один, а сколько угодно раз. Отметим, что повторный поджог можно осуществить не только от внешнего источника, но и пламенем, подошедшим по среде. Для этого достаточно линию фитилей, вдоль которой бежит пламя, замкнуть в кольцо, и пламя начнёт вращаться по ней [44].
Распространяющийся по аксону импульс также является автоволной; он представляет собой электрохимическую волну перехода между двумя состояниями: покоя, когда разность потенциалов на мембране волокна велика (приблизительно – 0.07 В), и активного состояния – возбуждения, когда разность потенциалов мала (около +0.02 В). При распространении нервного импульса в каждой точке возбудимой мембраны расходуется энергия, исходно запасённая в виде неравномерных концентраций ионов калия и натрия по обе стороны мембраны.
После прохождения нервного импульса мембрана остаётся деполяризованной и неспособной к следующему возбуждению (рефрактерный период). Чтобы восстановить возбудимость клетки, необходимо восстановить исходные потенциалы, то есть восстановить исходную концентрацию ионов (потенциал покоя).
Справится ли с этой задачей простой осмос? Ответ Ходжкина и Хаксли – нет! И тут на сцену выходят «молекулы помощники» – ионные насосы. Белковые молекулы, которые выкачивают из клетки ионы натрия и закачивают ионы калия. Причём делают это чрезвычайно быстро, судя по временному графику нервного импульса. Хотя, можно предположить, что ионные концентрации восстанавливаются за счёт их общего количества во внутри– и внеклеточном пространстве, а поддерживаются насосами.
Насосы
Мембранный электрический потенциал генерируется с помощью поддержания концентрации ионов, присутствующих в физиологических жидкостях организма и внутриклеточной среды.
Каждый потенциал действия оставляет клетку с бо́льшим, чем следовало бы быть, количеством натрия внутри и с бо́льшим количеством калия снаружи. Восстановить исходный баланс должен был бы осмос. Но нервные импульсы несутся по аксону один за другим с такой частотой, что медленный осмос не справится. И не забываем, что через мембраны ионы калия и натрия надо перемещать против градиента концентрации и электрохимического градиента. Ходжкин предположил, что этот дисбаланс исправляется особым белком, который транспортирует избыточные ионы натрия из клетки, а ионы калия – в клетку. В результате чего исходные градиенты концентраций натрия и калия восстанавливаются. [8]
1950-х годах Ходжкин обнаружил, что при возбуждении нерва расходуется АТФ, а также, что перенос катиона натрия из клетки замедляется, если подавить синтез АТФ. Начало развиваться представление о ферменте АТФазе, которую в тот момент считали ответственной за биосинтез АТФ.
Каждый потенциал действия оставляет клетку с бо́льшим, чем следовало бы быть, количеством натрия внутри и с бо́льшим количеством калия снаружи. Восстановить исходный баланс должен был бы осмос. Но нервные импульсы несутся по аксону один за другим с такой частотой, что медленный осмос не справится. И не забываем, что через мембраны ионы калия и натрия надо перемещать против градиента концентрации и электрохимического градиента. Ходжкин предположил, что этот дисбаланс исправляется особым белком, который транспортирует избыточные ионы натрия из клетки, а ионы калия – в клетку. В результате чего исходные градиенты концентраций натрия и калия восстанавливаются. [8]
В 1950-х годах Ходжкин обнаружил, что при возбуждении нерва расходуется АТФ, а также, что перенос катиона натрия из клетки замедляется, если подавить синтез АТФ. Начало развиваться представление о ферменте АТФазе, которую в тот момент считали ответственной за биосинтез АТФ.
Биохимия нейрона
Напомню, что белки – это полимеры – молекулярные «бусы», состоящие из «бусин» -мономерных аминокислот. Каждая аминокислота имеет: аминную группу, карбоксильную группу и радикал.
Всего в состав белков входят 20 типов аминокислот, которые различаются лишь радикалами. Самый простой из радикалов водород даёт нам аминокислоту, которая называется глицин.
Полимеризация аминокислот с образованием белка происходит за счёт связывания COOH-группы предыдущей аминокислоты с NH2 следующей (такая связь называется пептидной).
В результате появляются линейные цепочки, состоящие из сотен аминокислот (100 аминокислот уже белок, меньше ста ещё пептид).
Итоговая аминокислотная цепь – это первичная структура белка. Радикалы не принимают участия в её формировании. Средняя длина белка 300—700 аминокислот. У каждого белка своя уникальная структура, свой набор и порядок аминокислот.

Рисунок 38. Белок
Следующий этап – формирование вторичной структуры белка. Она происходит за счёт присутствия довольно больших зарядов внутри аминокислот: положительного на аминной группе и отрицательного на карбоксильной.
Под влиянием этих зарядов первичная структура начинает сворачиваться. Самый известный способ свёртывания – это спираль. На каждом витке такой спирали примерно три аминокислоты. Радикалы при этом вновь не участвуют.
На третьем этапе спираль сворачивается в белковый клубок. Его образование происходит за счёт взаимодействия радикалов. Они же все могут быть разными и положительными, и отрицательными. Именно в таком состоянии белок становится молекулярной белковой машиной. Теперь он способен работать, например, схватить какую-нибудь молекулу и что-нибудь с ней сделать.
Как это происходит. Благодаря своей химической структуре белок способен производить захват молекулы-мишени (лиганда), для каждого белка мишень своя. Белок подстраивается под свою мишень по принципу ключ-замок. После этого он способен выполнять с лигандом те или иные действия.
По типу операций с лигандом белки подразделяются на:
Белки-ферменты
транспортные белки
белки-каналы (насосы)
двигательные, защитные, строительные и др.
Как работает расщепляющий пищевой белок-фермент.
Захватить лиганд.
Разорвать его.
Отпустить.
А бывает наоборот – синтез новых веществ:
Захватить два лиганда.
Соединить их.
Отпустить.
Транспортный белок, например, гемоглобин работает так. Схватил кислород, перенёс его, отпустил и опять в лёгкие за новым кислородом.
В организме работает около 5000 групп ферментов.
Применительно к теме нервных клеток нам особенно интересны белки-каналы и белки-насосы.
Простейшие «открытые» белки—каналы условно представляют собой трубки, встроенные в мембрану клетки. через них может идти диффузия1 как правило строго определённых мелких частиц – молекул H2O, ионов K+, Na+ и прочих (то, что делает мембрану полупроницаемой и является основой для осмоса).
Большинство же каналов не такие простые, а со створкой – его отверстие перекрыто петлёй-створкой (канал закрыт). Но при определённых условиях створка может открываться, разрешая диффузию. Условия открытия могут быть разные – влияние химических веществ (гормонов), электрического потенциала или давления.
Работа белка-насоса:
1 Диффузия – движение частиц из области с более высокой концентрации в область с низкой концентрацией.
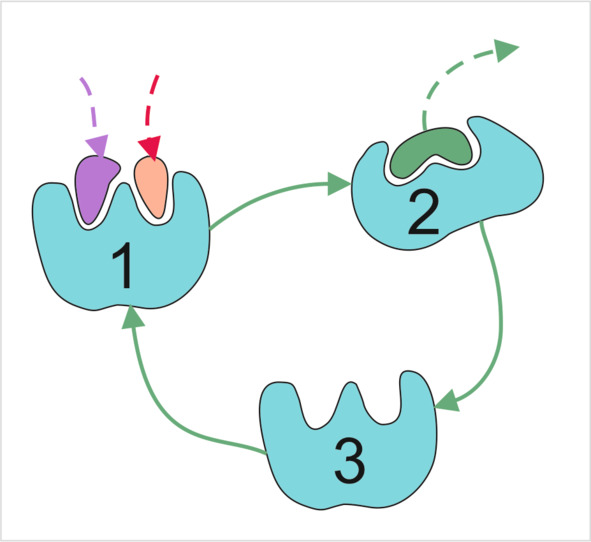
Рисунок 39 Работа белка-насоса
Есть ещё одна группа белков – это белки—рецепторы. Будучи встроенными в мембрану клетки (например, в месте синапса), они выполняют информационные функции. Лиганд в этом случае – сигнал об определённом событии во внешней межклеточной среде. Присоединяя лиганд белок-рецептор запускает ответную реакцию клетки влияя на её белки-каналы, насосы, ферменты.
Простейшие открытые каналы и белки—каналы со створкой являются пассивными элементами – они либо пропускают через себя микрочастицы, либо нет. А вот каналы-насосы способны выполнять работу (как и белки-ферменты, и белки-рецепторы). Такой насос захватывает лиганд с одной стороны мембраны и переносит его на другую. Такая работа уже требует затрат энергии. Для получения этой энергии белок умеет получать её от АТФ, отрывая от неё один или два остатка фосфорной кислоты.
Белки, потребляющие энергию АТФ для выполнения какой-либо работы, часто обобщённо называют – аденозинтрифосфата́зы (АТФ-азы).
АТФ
Молекула АТФ (аленозинтрифосфата) состоит из рибозы, аденина и трёх остатков фосфорной кислоты, между которыми имеются две высокоэнергетические связи. Энергия каждой из них составляет 30,6 кДж/моль. Поэтому её и называют макроэргической в отличие от простой связи, энергия которой составляет около 13 кДж/моль. При отщеплении от молекулы АТФ одного остатка образуется молекула АДФ (аденозиндифосфат), а при отщеплении двух остатков —соответственно молекула АМФ (аденозинмонофосфат).
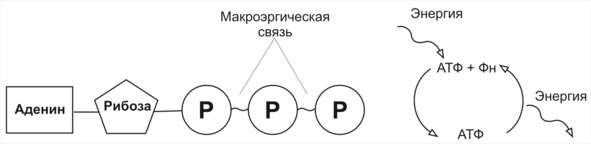
Рисунок 40 Строение молекулы аленозинтрифосфата (АТФ) и её роль в превращении энергии
Восстановление (синтез) молекул АТФ происходит в митохондриях, внутри самой клетки. Энергия запасается в результате реакций окисления органических веществ. Клетка использует эту запасённую энергию во всех процессах жизнедеятельности.
Насосы
Наиболее важными из белков-насосов, которые поддерживают мембранный потенциал, являются:
1. Натриево-кальциевый (транспортирует один ион Са2+ внутрь клетки в обмен на 3 иона Na+, транспортируемых наружу).
2. Натриево-калиевый (транспортирует один ион Na+ наружу в обмен на один ион К+ внутрь).
3. Хлорный (транспортирует из клетки наружу ионы Cl—).

Рисунок 41. Строение клеточной мембраны. Видны два слоя липидных молекул, ионные каналы и ионные насосы K+ и Na+.
Энергозатраты при возбуждении нейронов обусловлены главным образом работой натрий-калиевого насоса, который активируется поступлением внутрь протоплазмы ионов Na+.
В 1957 году Йенсом Скоу (дат. Jens Christian Skou 1918—2018) была открыта Na+/K+-АТФаза. Он выделил этот фермент из периферических нервов с помощью уабаина – специфически связывающегося с АТФазой гликозида. За это открытие спустя сорок лет, в 1997 году он был удостоен Нобелевской премии по химии.
Благодаря этому открытию был объяснён принцип работы «натрий-калиевого насоса» который поддерживает концентрацию ионов натрия в цитоплазме клетки на очень низком уровне по сравнению с внеклеточной средой.
Na+/K+-АТФ-аза (Na+/K+ аденозинтрифосфатаза) – фермент из группы транспортных аденозинтрифосфатаз, встречающийся в плазматической мембране всех клеток животных. Na+/K+-АТФ-аза переносит ионы К+ внутрь клетки, в то время как ионы Na+ выводятся наружу. Основная функция – поддержание потенциала покоя и регулирование клеточного объёма.
Как это работает. На первом этапе фермент присоединяет с внутренней стороны мембраны три иона Na+. Эти ионы изменяют конформацию1 третичной структуры белка. При этом фермент гидролизует одну молекулу АТФ. Выделившаяся в результате гидролиза энергия расходуется на проведение конформации белка, благодаря чему три иона Na+ и ион PO4 – (фосфат) оказываются на внешней стороне мембраны. Здесь ионы Na+ отщепляются, и присоединяются два иона К+. Далее фермент возвращается в исходную конформацию, а фосфат-ион и ионы К+ оказываются на внутренней стороне мембраны. Здесь ионы К+ отщепляются, и насос вновь готов к работе.
В итоге внутри клетки создаётся высокая концентрация K+, а во внеклеточной среде – высокая концентрация ионов Na+.
1 Конформа́ция молекулы – пространственное расположение атомов в молекуле определённой конфигурации, обусловленное поворотом вокруг одной или нескольких одинарных сигма-связей.
Математическая модель
В 1952 году для описания электрических механизмов, обусловливающих возникновение и распространение нервного сигнала в гигантском аксоне кальмара Аланом Ллойдом Ходжкином и Эндрю Хаксли разработана математическая модель, названная в честь авторов «Модель Ходжкина—Хаксли».
Точечная модель Ходжкина—Хаксли представляет собой систему обыкновенных дифференциальных уравнений, которая, в частности, пригодна для описания характеристик электрического сигнала.
Модель Ходжкина—Хаксли возникла не на пустом месте. Вот её предыстория.
Метод «интегрировать-и-сработать»
Одна из ранних математических моделей возбудимой клетки была предложена в 1907 году французским физиологом Луи Лапиком (Louis Lapicque, 1866—1952). Модель была описана следующей формулой:
,
которая есть производная по времени закона ёмкости, Q = CV. Если на вход системы подаётся ток, то разность потенциалов (напряжение Vm,) на мембране возрастает со временем, пока не достигает некоторого порогового значения Vth, при котором происходит скачкообразное изменение потенциала на выходе и напряжение сбрасывается до остаточного потенциала. После этого цикл работы повторяется с начала, пока опять не накопится энергия для следующего срабатывания. Эта модель имеет один существенный недостаток – бесконечно большое линейное возрастание частоты срабатывания при линейном увеличении входного тока, что возможно только в абсолютно идеальных условиях без утечек.
Поэтому модель была уточнена введением рефрактерного периода tref, который ограничивает частоту срабатывания, задерживая срабатывание на некоторое время после достижения потенциала действия. Частота срабатывания в этом случае может быть описана следующей формулой:
Недостаток этого подхода заключается в проявлении независимой от времени способности запоминания. Если модель получает некоторый заряд, недостаточный для срабатывания, она будет хранить его до следующего подзаряда. Если же дополнительного подзаряда не произойдёт – напряжение будет сохраняться вечно, что явно не соответствует процессам, наблюдаемым на реальной мембране.
Метод «интегрировать-и-сработать» с утечками
Исправить недостаток вечной памяти позволило введение концепции утечки. Метод моделирует имитацию диффузии ионов, происходящую в мембране в случае недостижения условий для генерации потенциала действия. Улучшенная подобным образом модель может быть описана следующей формулой:

где Rm – значение электрического сопротивления мембраны. Теперь, чтобы сгенерировался потенциал действия, значение тока на входе должно превысить некоторый порог Ith = Vth / Rm. Иначе происходит утечка, аннулируя любые изменения потенциала. Частота срабатывания принимает следующий вид:

что сходится с предыдущей моделью (без утечки) для больших величин тока.
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги