Глава II «Исход» древнейших представителей рода Homo из Африки как важный этап антропогенеза
Если последовательно придерживаться концепции моногенизма, следует признать, что факты, изложенные в первой главе, неизбежно наводят на мысль, что ранние этапы эволюционной истории человека когда-то должны были обусловить знаменательное событие в жизни наших далеких предков — выход за пределы границ африканской прародины. Достаточно сопоставить масштабы «колыбели» рода Homo с планетарными параметрами дисперсии этого таксона в течение последующих эпох. Для того далекого времени «исход» человека из Африки вполне можно сопоставить по значению с выходом в космос в двадцатом веке.
В данной главе будут рассмотрены представления, сложившиеся в современной антропологии и археологии по вопросам «где?» и «когда?» произошло описываемое событие, которое можно считать conditio sine qua поп для становления всего человечества нашей планеты и Разума в нашей части Вселенной. Впрочем, оно, как и любой другой крупный феномен, было подготовлено предшествовавшим ходом развития и не могло не произойти в силу накопленного потенциала универсальных адаптаций и видового опыта внутриафриканской дисперсии. Комплекс приспособлений, выработанных в африканской саванне, явился преадаптацией для более сложных реакций на разнообразные средовые воздействия и обусловил возможность новых действий таксона для освоения безлюдных земель. Конечно, не менее важным является вопрос «кто?» — точнее какой вид рода Homo впервые «перешагнул» африканский географический рубеж и вышел на просторь Евразии.
Первоначальное юявление древних Homo на внеафриканских территориях будет темой следующих разделов работы. В данной главе внимание уделяется именно моменту выхода каких-то плио-плейстоценовых гоминид за пределы Африки, так сказать «акту пересечения границы» континента. При этом, естественно, можно говорить о месте и времени описываемого события лишь в порядке более или менее вероятного научно обоснованного предположения.
Прежде чем перейти к поставленным выше вопросам «кто?», «где?» и «когда?» необходимо остановиться на широко обсуждаемой в научной литературе проблеме возможных причин исхода гоминин из Африки, т. е. практически ввести в обсуждение еще один вопрос — «почему?». Иссле
— дователи миграций млекопитающих делят последних на две контрастные категории — широко распространяющихся видов (widely dispersing species) и не распространяющихся видов (non dispersing species). Человеческий род у своих истоков отличался ограниченной дисперсией, подобной тому, что мы наблюдаем у человекообразных обезьян, но примерно 1,8 млн. лет назад приобрел тип дисперсии широко распространяющихся крупных млекопитающих (Antón et al., 2000). Имеется мнение, согласно которому приобретение древними гоминидами возможности выйти за пределы Африки было обусловлено технологическим прогрессом, связанным с развитием ашельской традиции в технологии изготовления орудий (Wolpoff, 1999, after Antón et al., 2000). Эта точка зрения была рассмотрена в ряде специальных работ, в частности — в упомянутой выше статье (Antón et al., 2000) и вызвала серьезную критику. Прежде всего нельзя недооценивать те факты, что известные древнейшие миграции Homo (Дманиси, Лонггупо) принесли с собой не ашельскую технику, а изделия типа Mode 1 (либо вообще никакого каменного инвентаря) и распространение самых ранних Homo в Восточной и Юго-Восточной Азии не повлекло за собой экспансию техники Mode
Концепция «линии Мовиуса», пока не утратившая полностью своего значения, является одной из наглядных иллюстраций отсутствия прямой причинной связи между ашелем и древними миграциями гоминид. Более того, даже значительно более поздние местонахождения (Чепрано) также не включают орудий ашельского типа. Отсюда делается вывод (ор. cit.), что переход на ашельскую традицию не был основным стимулом миграций древнейших представителей Homo. Можно добавить к этому, что крупные млекопитающие (бовиды, эквиды, крупные хищники), разумеется, никак не связанные с какой-либо технологией, свободно перемещались между Африкой и Азией в описываемые периоды времени (Kurten, 1968, cit. after Antón et al., 2000). Необходимо при всем этом отметить, что в работах М. Волпоффа в связи с анализом роли прогресса технологии как стимула миграций, подчеркивается важная роль развития когнитивных способностей у гоминид в ходе расселения. К тому же сводится замечание А. Тернера (1997) о роли эволюции мозга в таких процессах. Эти мысли несомненно затрагивают очень важный аспект теории антропогенеза, включая проблемы плио-плейстоценовых миграций гоминид.
При рассмотрении причин, обусловивших «исход» древних групп Homo из Африки часто затрагивается вопрос о значении экологических факторов. Так, например, американский исследователь О. Бар-Йозеф в работе 1997 года пишет, что «первые миграции гоминид из Африки были вызваны изменениями климата, которые произошли около 1,6 млн. лет назад» (стр.56), однако на карте (рис. 2, стр. 42) в этой работе автор предпочитает назвать несколько более раннюю дату «исхода» из Африки — 1,8 млн. лет. Оставив пока в стороне предполагаемую датировку описываемых событий, отметим, что само по себе влияние резких климатических изменений глобального и, в частности, африканского масштаба, на все стороны жизни гоминид не вызывает сомнений. Требует специального рассмотрения вопрос о механизме и характере воздействия этого фактора на процессы видообразования и миграций. Поведение плио-плейстоценовых гоминид следует рассматривать в более широких рамках закономерностей «экологии и эволюции наземной фауны млекопитающих» (Тернер,

Схема предполагаемых путей первоначального расселения гоминин из Африки. По работам нескольких авторов: О. Бар-Йозеф (1997) (непрерывная линия), С. Миттен и М. Рид (2003) (пунктир).
1997). С этой формулировкой нельзя не согласиться, особенно если речь идет о столь древних периодах времени, однако даже тогда гоминиды имели свои особенности контакта с окружающей средой, которые выделяли только что возникший род Homo из общей массы таксонов, захваченных единым потоком радикальных изменений биосферы.
Принимая наиболее распространенную точку зрения, согласно которой «исход» имел место в конце периода «колыбели» на эволюционной стадии Homo erectus, авторы публикаций на рассматриваемую тему называют несколько вероятных причин расселения ранних Homo за пределы Африки. Так, М. Л. Бутовская (1997) считает, что такими причинами были: общий рост численности населения (этот фактор приводится в первую очередь), в основе которого, по мнению автора, лежит увеличение продолжительности жизни, совершенствование стратегии заботы о детях, развитие альтруизма и, далее, — прогресс методов добычи и обработки пищи (освоение огня), увеличение размеров территорий, занимаемых различными группами, способность варьировать пищевые стратегии (Бутовская, 1997, стр.25). Последний тезис я связал бы непосредственно с произошедшим у истоков рода Homo переходом к всеядности. Кстати, такая резкая и быстрая перемена состава пищи и, соответственно, пищевой стратегии, иногда кажется мало вероятной, однако можно на это возразить, что в эволюции ранних гоминид уже имелись определенные предпосылки пищевой деспециализации. Так, существует мнение, что отдельные виды австралопитеков, включая древних, обладали значительной степенью универсализма в этом отношении. В частности, A.africanus, которого некоторые исследователи считают близким к эволюционной линии человека (и даже предком последнего), «…was highly opportunistic and adaptable in its feeding habits» (…«был в высшей степени гибким и адаптабельным в своей пищевой стратегии» [van der Merwe et al., 2003, p. 581]).
М. Волпофф в работе, озаглавленной «Из Африки» (1997) пришел к выводу, что «Люди ушли из Африки не по принуждению и не из-за голода. Это была благоприятная возможность использовать развившиеся адаптационные возможности с максимальной выгодой для себя. Необходимые предпосылки сформировались в результате менявшихся анатомических адаптаций к дневной активности, включавших большие размеры тела, длинные ноги и ряд физиологических изменений, а также благодаря использованию сложных орудий» (Волпофф, 1997, стр. 38). Говоря об орудийном факторе расселения рода Homo, автор имеет в виду более общую и глубокую причину экспансии этого рода — прогрессирующее совершенствование мозга, объем которого постоянно увеличивался в процессе эволюции на стадии «колыбели» и уже на поздних этапах существования Homo rudolfensis превысил 770 см3.
Рассматривая причины «исхода» в оценке разных авторов, можно сделать несколько обобщающих заключений, вытекающих из сравнительного анализа высказываний по данной теме.
Констатируется иерархический характер системы предполагаемых факторов, которые стимулировали движение популяций первых Homo в направлении «Out-of-Africa». В качестве основного «макро-фактора» называют глобальное изменение климата, начавшееся еще в конце миоцена и достигшее ко времени 2,5 млн. лет до современности практически масштабов экологической катастрофы. Этот фактор включал похолодание, аридизацию, формирование обширных открытых пространств вымирание многих видов (в частности — гоминид), а для рода Homo означал необходимость существенной перестройки всего морфофизиологического и поведенческого комплексов, присущих предковым формам — австралопитековым: увеличение общих размеров тела, удлинение ног в связи с жизненно важной задачей преодоления больших расстояний с периодическим переходом на быстрый бег (новый этап эволюции бипедии), совершенствование механизмов терморегуляции. Большое значение для эволюционирующего рода Homo имел переход на животную, более калорийную пищу и выработка стратегии «активного скэвинджинга». Количественное и качественное (что особенно важно) совершенствование диеты способствовало ускорению эволюции в ее специфическом человеческом варианте, а новая стратегия добычи пищи приводила к необходимости постоянных длительных переходов, улучшению организации групп, совершенствованию орудий. Это лежало в основе значительного и быстрого расширения территории, занимаемой древнейшим (или древнейшими) видом (видами?) рода Homo. Исследование, проведенное антропологами (Antón et al., 2002), показало, что площадь территории (HR), освоенной видом Homo erectus, превышала HR австралопитековых в 10 раз (!), что вызвало экспансию вида, ставшую уже жизненно необходимой для выживания. Все эти аспекты прогресса вместе взятые означали формирование нового вида, которое я отнес бы к системе макро-факторов, явившейся предпосылкой и стимулом «исхода» гоминид из Африки.
Рассматривая значение процессов видообразования в ходе формирования поведенческого комплекса Homo как «широко расселяющегося вида», необходимо прежде всего отметить континентальный масштаб внутриафриканских миграций в конце периода «колыбели». Если мы стали бы игнорировать этот важнейший этап эволюционной истории рода Homo, мы могли бы впасть в серьезную ошибку, предположив, что описанный выше прогрессивный комплекс, сложившийся в пределах этого рода, не был реализован до момента «исхода». Палеоантропологические находки, относящиеся к периоду 1,7–1,8 млн. лет до современности на юге и северо-западе Африки свидетельствуют о значительно возросшей миграционной активности гоминид, отражающей большие возможности формирующегося рода Homo в отношении проникновения в новые регионы с перспективой их колонизации. Здесь можно выделить еще один достаточно хорошо аргументированный фактор, явившийся стимулом «исхода»-Homo из Африки: практический опыт ранних человеческих миграций в освоении обширных территорий в пределах тропического и субтропического пояса, накопленный в Африке на базе морфофизиологических и психологических характеристик эволюционирующего нового вида. Добавим к этому еще один макро-фактор, на этот раз — географический по своей природе, — определивший описываемое событие в жизни гоминид: наличие смежных с Африкой территорий со сходными природными условиями, являющихся как бы продолжением знакомых, хорошо освоенных африканских земель.
О миграциях этого времени более или менее обоснованно мы можем судить в основном по археологическим материалам. Относительно видового состава групп, впервые вышедших за пределы Африканского континента, известно пока недостаточно. Их физический тип устанавливается по самым ранним экстра-африканским находкам, представленным скелетными остатками из Дманиси, а также индонезийскими формами. Большинство антропологов считает, что «исход» из Африки был осуществлен гомининами, относящимися к виду Homo erectus (Бар-Йозеф, 1997; Mithen & Reed, 2002), однако не все специалисты с этим согласны. Так, Р. Деннелл в своей работе, посвященной миграциям гоминид (Dennell, 2003), признавая, что датировка древнейших индонезийских находок (1,8 млн. лет) позволяет говорить об очень раннем «исходе» из Африки, в то же время высказывает сомнение относительно интерпретации факта достижения Явы гоминидами этого времени как основы для суждения о первом выходе человека из Африки. Р. Деннелл подразумевает возможность нескольких «исходов» из Африки в разное время, включая более ранние, и тем самым ставит проблему биологических видов гоминид, начавших период расселения за пределами Африки. Эти вопросы интересуют многих специалистов, разрабатывающих гипотезы первоначального заселения Евразии и считающих, что «способность человечества к расселению проявилась очень рано, гораздо раньше, чем было принято думать» (Peretto, 2000, p. 175). Итальянский археолог К. Перетто, из работы которого взята приведенная выше цитата, приходит к логичному для такой теоретической позиции выводу, что можно выдвинуть гипотезу распространения из Африки более древних форм, чем Homo erectus. Другой итальянский исследователь — Дж. Манци также считает неправильным ограничивать возможное решение вопроса о видовом составе группы (или групп) «исхода» из Африки предположением, что первым покинул пределы Африки только Homo erectus/ergaster и вносит осторожное уточнение: «Ното ergaster или другой, более архаичный вид африканского происхождения» (Manzi, 2000, p. 117). Нетрудно на основании современных данных понять, что речь в этом контексте может идти о Homo habilis sensu lato, т. е., — если встать на позицию сторонников гипотезы двух видов первых Homo, — Н. habilis sensu stricto или Н. rudolfensis. Археологические материалы не противоречат такому решению: практически можно утверждать, что все или почти все каменные орудия, относящиеся к древнейшим местонахождениям гоминид вне Африки, могут быть определены как Mode I (олдован), т. е. к доашельской стадии. К сожалению, эти данные не позволяют судить о физическом типе изготовителей орудий, т. к. Mode I может сопровождать скелетные остатки как Homo erectus/ergaster, так и Homo rudolfensis. Так, например, артефакты типа олдован найдены вместе с черепом из Чепрано, краниологический комплекс которого бесспорно свидетельствует о принадлежности к Homo erectus (sensu lato) и тот же Mode I характерен для древнейших африканских форм Homo habilis/ rudolfensis.
Принципиальная возможность выхода за пределы Африки пре-эректоидных гоминид нуждается в конкретных антропологических доказательствах. Решающими данными в этом отношении могли бы быть прежде всего материалы из Дманиси и находки Homo erectus mojokertensis из Индонезии.
Раскопки в Дманиси (Грузия), начиная с 1991 г., дали важнейший материал по ранним этапам эволюции рода Homo, относящийся к нижне-виллафранкскому времени (1,77 млн. лет до современности). Обнаружено 4 черепа разной сохранности и фрагменты посткраниального скелета: calvarium D2280, часть черепа D2282, соединяемая теперь с нижней челюстью D211, череп 2700/2735, а также нижняя челюсть D2600. Исследователи отмечают на найденных черепах слабое развитие надглазничного валика и других суперструктур, сильную выраженность посторбитальной констрикции, расширение черепов в области сосцевидных отростков при округлой общей форме затылочного отдела, тенденцию к «килевидной» форме черепной крышки, выступание средней части лицевого отдела (Rightmier et al., 2006). Удалось получить довольно представительный материал по морфологии зубов в серии Дманиси. В цитируемой работе даются размерные характеристики верхних зубов в черепе D2700 и нижних зубов в челюсти D2735. Следует отметить своеобразную форму верхних резцов, коронки которых резко расширяются к режущему краю (MDcor=12,6 мм) и крупные размеры моляров, причем первый слегка превосходит второй по мезиодистальноиу диаметру и почти равен ему по вестибулолингвальному: MDM1=13,0 mm, VLM1=13,0 mm; MDM2=12,6 mm, VLM2=12,9 мм (приведены средние величины правой и левой сторон, которые, впрочем, практически не различаются, кроме одного случая, когда VL правого М2 в точности равен VLM1). Коронки первого и второго верхних моляров в поперечном сечении имеют почти квадратную форму, что является архаичной чертой, главным образом в отношении второго моляра, еще не затронутого редукционным процессом. К числу примитивных особенностей можно также отнести почти параллельное расположение рядов моляров и премоляров и довольно крупный клык. Зато нужно особо обратить внимание на редукцию третьих моляров, в наибольшей мере по мезио-дистальному диаметру. Здесь мы имеем дело с существенным эволюционным сдвигом в сапиентном направлении, что можно сказать и относительно превосходства первого моляра над вторым по размерам коронки. Авторы рассматриваемой статьи делают вывод, что по размерам первых двух верхних моляров экземпляры из Дманиси близки к размерам аналогичных зубов африканских Homo erectus.
На нижней челюсти D2735, как и на верхней, можно констатировать наличие некоторых прогрессивных особенностей. Намечается редукция пятого бугорка — гипоконулида — на обоих нижних молярах. Foramen mentale располагается под первым премоляром, как у современного человека. Однако, при всем этом морфологический облик зубов описываемой челюсти выглядит в целом архаично. Клык по масштабу рода Homo крупный, имеется небольшая «филогенетическая» диастема между клыком и премоляром с обеих сторон челюсти, премоляры и моляры крупные: MDP3=9,5 мм, VLP3=10,5 мм; MDP4=7,4 мм (средняя между правым и левым зубом), VLP4=9,4 мм. Для сравнения приведем размеры нижних премоляров Homo erectus по Ф. Вейденрейху: Р3 — 8,8×9,9; Р4 — 9,0×9,8 (Weidenreich, 1937). Первый премоляр в челюсти D2735 заметно превышает величину того же зуба у Homo erectus pekinensis. Что касается величины второго премоляра, то приведенная авторами цифра 7,4 мм для MD чрезвычайно мала для Н. erectus и отражает (если это не ошибка) особое своеобразие серии из Дманиси. Вестибулолингвальный диаметр второго премоляра несколько меньше, чем у «синантропа». Нужно заметить, что этот зуб в челюсти D2735 имеет очень характерные для древних гоминид черты строения: резко выступающий дисто-лингвальный угол коронки в связи экспансией талонида и мезиальный сдвиг вершины метаконида. Эти особенности придают коронкам вторых нижних премоляров из Дманиси форму искаженного прямоугольника, которая была отмечена на ископаемых среднеплейстоценовых зубах прежними исследователями (Martinon-Torres et al., 2005; Bailey & Lynch, 2005).
Нижние моляры в челюсти D2735, как и верхние, характеризуются мозаикой эволюционно неравноценных морфологических особенностей: третий моляр редуцирован, что видно по ячейке правого М3 (левый отсутствует). Первый моляр практически равен второму по мезиодистальному диаметру (MDM1=13,1 мм, MDM2=13,1/13,5 мм) и существенно больше по вестибулолингвальному (VLM1= 11,4 мм, VLM2=10,4 мм). Мезиодистальный диаметр первого моляра D2735 значительно превосходит MDM1 синантропа (12,6 мм), а вестибулолингвальный по сравнению с Н. erectus несколько меньше (синантроп — 11,8 мм). Второй нижний моляр в рассматриваемой челюсти по MD существенно больше, чем у Н. erectus (12,6 мм), а по VL — гораздо меньше (синантроп — 12,5 мм). Приведенные выше размеры нижних моляров D2735 позволяют заключить, что эти зубы имеют своеобразную форму контура коронок, что еще более ярко проявляется в величинах индекса VL/MD. Индекс первого моляра равен 86,7 (средняя между правой и левой сторонами), второго — 78,2. Эти цифры отражают значительную степень вытянутости коронок моляров в мезиодистальном направлении. На зубах Homo erectus pekinen-sis соответствующие показатели равны 93,7 и 99,2, а у современного человека 92,9 и 92,7. Такие низкие индексы коронок нижних моляров могли бы показаться уникальными, если бы не данные по первому нижнему моляру Н. rudolfensis из Начукуи (Кения), индекс коронки которого равен 84,7 (Prat et al., 2005). Вероятно, эта особенность была в какой-то мере связана с ранними Homo. Индексы, полученные для серии зубов дрёвнейших людей Африки в среднем не позволяют определенно подтвердить этот вывод, однако минимальные величины, — 82,8 и 85,6 (Brauer & Schultz, 1996) — свидетельствуют о том, что удлиненная форма нижних моляров у людей «эпохи колыбели» встречалась, хотя мы сейчас не можем сказать насколько часто. Во всяком случае, эти факты говорят о значительном размахе изменчивости ранних Homo. По данным тех же авторов в более поздние эпохи существования человека (ранний и поздний Н. erectus) значения описываемого индекса существенно повышаются, а изменчивость уменьшается.
В морфологии нижних моляров D2735, особенно — второго, бросается в глаза широкий и высокий гребень эпикристид, средний гребень тригонида, отделяющий глубокую переднюю ямку — fovea anterior. Эта структура в целом полностью сохранилась на эволюционной линии, завершившейся неандертальским таксоном и стала уникальной особенностью нижних моляров неандертальцев (Зубов, 1968; Зубов и Халдеева, 1989. Bailey,2006). У Н. sapiens она практически не встречается, у Homo hei-delbergensis редка. На нижних молярах D2735 хорошо просматриваются также задние ямки. Относительно велика ширина тригонида — признак, отмеченный Ф. Вейденрейхом на зубах синанатропа и охарактеризованный как примитивная особенность.
Для основной темы данной работы наибольший интерес представляет оценка объема черепов, давшая сенсационные результаты: 600–775 см3 (!), что хорошо укладывается в рамки показателей, типичных для Homo habilis/rudolfensis, хотя согласно предварительному таксономическому диагнозу дманисская серия была отнесена к Homo erectus. Интересно, что максимальный объем, зафиксированный в Дманиси, в точности совпадает с объемом наиболее известного «типового» черепа Homo rudolfensis — экземпляра KNM-ER 1470 из Кении. Сходство не ограничивается размерами: умеренно развитый надглазничный валик, округлый затылок, вогнутый контур лицевого отдела в латеральной норме, выраженный альвеолярный прогнатизм (рис. 3). Авторы опубликованной недавно детальной работы, посвященной черепам из Дманиси (Rightmier et al., 2006) недаром включили в число сравнительных экземпляров ряд черепов Homo habilis/rudolfensis (ОН7, ОН13, ОН24, KNM-ER 1470, KNM-ER 1813), причем обнаружено значительное сходство и констатируется чрезвычайно архаичный краниологический комплекс находок из Дманиси: «…there are resemblances to H. habilis (sensu stricto). Some of the characters considered seem to be primitive, in the sense that they are common not only to early Homo but also species of Australopithecus and extant apes» (op. cit. p. 136). («…имеются черты сходства с Homo habilis (sensu stricto). Некоторые из рассматриваемых особенностей представляются примитивными в том смысле, что они обычны не только для ранних Homo, но также для некоторых вцдов австралопитеков и современных обезьян». Однако, следует и противоположное по смыслу заключение:

Сравнение черепов KNM-ER 1470 (справа) по работе: D. Johanson & В. Blake «From Lucy to language», 1996 и Дманиси D 2700 (слева) по работе: G. Ph. Rightmier, D. Lordkipanidze, A. Vekua «Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia». Journal of Human Evolution. 2006. N 50. P. 115–141
«…but reveal numerous resemblances to H.erectus» («…но обнаруживают многочисленные черты сходства с Homo erectus») (там же), сделанное на основе сравнения с экземплярами KNM-ER3733, KNM-WT 15000, KNM-ER 992. Отмечено также сходство с черепами из Индонезии (фауна Джетис) — Сангиран 4 и ребенок из Моджокерто (древность соответственно 1,66 и 1,81 млн. лет до современности). Авторы рассматриваемой работы склонились к признанию большей близости с Н. erectus, но весь ход анализа показывает, что выбор между двумя видами ископаемых гоминид представлял существенные трудности и окончательный таксономический диагноз в известной мере остается условным. Нам представляется более удачной острожная формулировка, данная авторами в резюме рассматриваемой работы: «Проведенное исследование показывает, что находки в Дманиси близки к стволу, от которого эволюционировал H. erectus» («close to the stem from which H. erectus evolved» (op. cit., p. 115). Описанная работа, на наш взгляд, убедительно показала, что в Дманиси найдена промежуточная форма эволюционирующего таксона внутри рода Homo, дающая основание для вывода о физическом облике гоминид, осуществивших «исход» из Африки в ранне-виллафранкское время. С нашей точки зрения логично предположить, что наука близко подошла к решению проблемы антропологического типа и эволюционного уровня гоминид, впервые покинувших пределы Африканской прародины рода Homo. Это был промежуточный, формирующийся таксон, сохранивший черты анатомических комплексов Homo habilis sensu Iato, но обладающий уже набором морфофизиологических и поведенческих адаптивных особенностей, обеспечивавшим переход к радикальной перестройке образа жизни и подъем на новую, более высокую эволюционную ступень. Фактически это еще не был Н. erectus, но уже трудно отнести его безоговорочно к Н. habilis. В этом случае следовало бы говорить о подвиде, притом достаточно хорошо очерченном, чтобы считаться самостоятельным таксоном в цепи эволюции рода Homo и имеющем право на собственное наименование в систематике гоминин.
В связи с этим необходимо подробнее рассмотреть вопрос о том, каково может быть это наименование и какой биномен соответствовал бы эволюционному уровню и систематическому положению исследуемой формы Homo. Казалось бы, проблема не очень сложна — надо только строго соблюдать правила биологической систематики. Однако, когда речь идет о человеке, притом в аспекте вертикальной систематики эволюционирующих и мигрирующих форм, возникает ряд трудностей, на которые антропологи не раз обращали внимание в подобных ситуациях и, в частности, в данном конкретном случае, когда речь шла о самых ранних представителях человеческого рода.
Процесс видообразования у человека в той или иной мере всегда отличался практически неповторимой спецификой. В моих прежних работах (Зубов, 1982, 1985, 2004) неоднократно обосновывалась точка зрения, согласно которой для антропогенеза характерен феномен «сдерживания адаптивной радиации», за счет особой формы эволюции, — магистрализации, сущностной чертой которой является акцент отбора на развитие мозга и, соответственно, — на сложные формы поведения. При этом частично отпадает и, во всяком случае, уменьшается зависимость от окружающей среды и необходимость значительных морфофизиологических адаптивных перестроек, а значит — ослабевает тенденция видообразования, которой противостоит стихийная видовая «таксономическая интеграция». Развитие материальной культуры, совершенствование орудий, накопление информации и совершенствование процессов обработки ее эволюционирующим мозгом составило базис «автономизации» (эмансипации) ветви Homo, которая, в конечном счете, несмотря на широкое расселение по всей планете, в систематическом отношении сконцентрировалась в таксономическом пространстве одного вида. Внутривидовая изменчивость не уменьшается и может возрастать, вследствие чего таксономический вес размаха вариаций соответственно падает. В этом состоит специфически «человеческий» феномен, называемый мною «интегрированным многообразием» (Зубов, 2004). Это свойство сохранять видовую модель, как бы уклоняясь от специализации, я назвал эффектом «избегания оптимумов» гармонии со средой, создающей стрессовые условия как основу возможной специализации. У человека в процессе антропогенеза ответом на видообразующий (и опасный для популяции) стресс была не тотальная морфофизиологическая перестройка, а движение, миграция, как поиск более подходящих условий жизни и уход от дистресса. Кстати, это было одной из побудительных причин больших перемещений групп древних Homo, таких, как рассматриваемая здесь «Великая миграция».
К сходным выводам относительно особенностей видообразования рода Homo, пришел и американский биолог А. Кларк Аркади считающий, что поведение может в значительной степени влиять на тенденции к специализации: представители рода Homo обладали высоким уровнем толерантности по отношению к среде обитания и в малой степени были склонны к специализации. Это свойство у плейстоценовых гоминид проявлялось в своего рода «упругости», «эластичности» таксона («resilience»), в ответ на деформирующее действие среды в период миграций (Arcadi, 2006).
Изложенные выше точки зрения приводят к заключению, что в пределах рода Homo видообразование не было очень интенсивным и «веер» адаптивной радиации был довольно ограниченным даже в период больших миграций. Однако, существуют и противоположные мнения. Так, И. Тэттерсол полагает, что антропологи всегда недооценивают число отдельных видов в ископаемом материале и что «преувеличив видовое разнообразие древнего таксона, мы, по крайней мере, сделаем меньше ошибок, чем в случае его преуменьшения» (Tattersall, 1986 after Manzi, 2000, p. 120). Индийские генетики В. Эсваран с коллегами (Eswaran et al., 2005) считают, что при расселении таксона из единого центра (в данном случае — Африки) он не мог сохранить исходный генотип в силу «диффузии демов, метисации и естественного отбора». Заметим, что речь в работе индийских исследователей идет о расселении Homo sapiens, который на пути своих миграций встречал различные группы архаичных гоминин и потому метисация действительно могла сильно модифицировать исходные генотипы, которые, по данным цитируемых авторов, могли ассимилировать до 80 % «не-африканского» генного материала. Однако, в отношении «Великой миграции» вопрос о метисации не имеет столь принципиального значения, т. к. все демы мигрирующего таксона были генотипически близки и метисация между ними, вероятно, мало влияла на общий генофонд потока миграции. Конечно, замечания относительно роли естественного отбора и мутаций справедливы, но, как говорилось выше, «эволюционная пластичность», свойственная человеческому роду, противостояла видообразующему действию изоляции и снижала возможность адаптивной радиации и специализации. Говоря о роли отбора, нельзя забывать о его стабилизирующей составляющей, которая должна была сохранять выгоды родовой модели Homo, отбрасывая нежелательные, притом менее значительные модификации, снижающие выживаемость магистрального таксона.
В результате анализа описанной дискуссии мы приходим к выводу, что в процессе «Великой миграции», которая, естественно, несла в себе известный «заряд» видообразования, таксономическая дифференциация обычно не превышала подвидового уровня. При этом и в подобных случаях таксономическая диагностика может быть субъективной по той причине, что «таксономическая значимость морфологических и генетических различий между близкими формами часто неясна» (Tattersall, 1986; Jolly, 1993; Mayr, 2000; Ahern et al., 2005, after A. Clark Arcadi, 2006, p. 1).
Сделав приведенные выше общие выводы относительно эволюционной и таксономической сущности промежуточных форм ранних Homo, перейдем теперь к конкретному анализу систематического статуса и выбору соответствующего биномена для той формы эволюционирующих гоминин, которая, с нашей точки зрения стала основой древнейшей волны миграции «out-of-Africa». Прежде всего необходимо оценить современное состояние этой проблемы, естественно вызвавшей поток литературы и острые, до сих пор не разрешенные противоречия. В 1977 году антропологи Н. Т. Боас и Ф. К. Хауэлл опубликовали интересное исследование фрагмента черепа с зубами из верхних слоев формации Шунгура (Эфиопия) (Boaz & Howell, 1977).
Датировка находки — 1,84 млн. лет (калий-аргон). Авторы считают ее промежуточной формой между Homo erectus и Homo habilis. Проводятся параллели с ОН24, ОН13 и Сангиран 4 и делается вывод, что найдена часть черепа раннего представителя Homo обитавшего в саванне. Было произведено измерение зубов (Р3-М3 d). Размеры очень близки к Н. habilis, больше, чем у синантропа и меньше, чем у массивных австралопитеков. В описываемой работе практически впервые специально уделяется значительное внимание таксономическому статусу и наименованию промежуточных форм между Н. habilis et Н. erectus. Используются материалы по древнейшим азиатским формам Meganthropus paleojava-nicus и Homo modjokertensis (терминология, данная в статье), которые по данным Ф. Тобайаса и Г. Кенигсвальда, сходны с ОН7 и ОН13 (Н. habilis), употребляя при этом удачный, во всяком случае на первый взгляд, термин «pre-erectus». Далее описание переносится на Африканский континент, где авторы также выделяют формы, которые можно отнести к «пре-эректусам», а именно: фрагментарные черепа SK847 et SK45 (Сварткранс) и некоторые находки в Олдувайском ущеслье и Ко-оби Фора (древность 1,8 млн. лет). Н.Боас и Ф. Хауэлл заключают, что в тропическом поясе Старого Света обнаруживается все больше следов «пре-эректусов», живших в Африке и Азии в период 2,3–1,5 млн. лет. Важным итогом рассматриваемой статьи является выделение отдельного таксона, промежуточного между хабилисом и эректусом. Пользуясь термином «pre-erectus», авторы, в то же время, не хотят закрепить его за описанной формой ввиду того, что это было бы нарушением правила приоритета, принятого в биологической систематике и предпочитают сохранить за выделенным таксоном (который они считают видом) первое название Homo mojokertensis.
Признавая правомерными некоторые рассуждения авторов рассматриваемой работы, я, в то же время, не считаю возможным согласиться полностью с таксономической оценкой «пре-эректуса» по ряду нижеследующих причин. Термин «mojokertensis» слишком однозначно «привязан» к месту обитания, слишком «географичен» и не отражает эволюционной сущности таксона, базируясь лишь на географической изменчивости. В 70-е годы еще не было столь важных материалов, как Дманиси, а без них морфологическая и таксономическая характеристика «пре-эректусов» теперь представляется невозможной, и вряд ли целесообразно считать «типовой» группой для «пре-эректусов» индонезийские формы, тем более, что «собственно моджокертские» находки фрагментарны, а их отношение к более представительной серии из Дманиси проблематично («Similarities to specimens from Sangiran are not numerous» («Черты сходства с экземплярами из Сангирана немногочисленны…» [Rightmire et al., 2006, p. 139], хотя что-то общее все же имеется.
Для палеодема Дманиси предложен триномен Homo erectus georgicus с оговоркой, что принадлежность массивной челюсти D2600 пока является предметом спора между исследователями, обнаружившими и описавшими эти ископаемые материалы. Относительно этого термина можно высказать те же сомнения, которые рассмотрены выше в связи с определением mojokertensis — узко локальный географический подтекст названия, которое должно охватывать огромный ареал и, что самое главное, отражать эволюционный статус подразделений рода Homo.
Казалось бы, адекватным для рассматриваемого таксона было бы название Н. pre-erectus, но оно не может быть преобразовано в триномен: термин «Homo erectus pre-erectus» включал бы противоречие внутри себя (подвид исключает вид). Мне же представляется, что рассмариваемая форма имела подвидовой статус.
Название описываемой эволюционной стадии должно соответствовать по своему содержанию, понятию «ранний Homo erectus». Остается только признать Н. pre-erectus видом, не входившим в состав Н. erectus, но находившимся «на пороге» эректуса, либо подвидом вида — предшественника, т. е. H. habilis pre-erectus. Мне представляется, что оба термина правомерны, но я все же склонен был бы предпочесть триномен Homo habilis pre-erectus, хотя он противоречит практически всеобщему убеждению, что первой мигрирующей формой человека был Н. erectus. Считаю здесь уместным поблагодарить д. и.н. С. В. Васильева за ценные замечания, высказанные им в процессе дискуссии по данному вопросу. Достоинством предлагаемого триномена является, с моей точки зрения, сохранение термина «pre-erectus» и статуса подвида за этой промежуточной формой. Наличие у пре-эректуса бесспорных черт хабилиса не раз подчеркивалось современными исследователями (Rightmier et al., 2006).
Данное предложение, как впрочем и почти любое другое, может быть объектом критики, хотя бы потому, что по правилам систематики следует отдавать предпочтение ранее присвоенным наименованиям. Однако, если мы обратимся к истории систематики гоминид, то практически на каждом шагу встретим примеры нарушения этого правила в связи с появлением новых находок, новых теоретических концепций, а также с постоянными спорами о таксономическом ранге тех или иных ископаемых групп. В ходе пересмотра систематики рода Homo были забыты синантропы, атлантропы, африкантропы, телантропы, что не было следствием случайных ошибок, а явилось необходимым отражением прогресса науки о человеке. Можно быть уверенным, что и дальше подобная ревизия систематики и номенклатуры гоминид будет время от времени неизбежна, в связи с чем я считаю правомерными новые и новые предложения о наименовании и таксономическом статусе предков человека, в частности — сформулированное выше рассуждение по поводу места «пре-эректуса» на эволюционном древе человека. Конечно, в подобных случаях необходимо обстоятельное обсуждение проблемы с привлечением новых фактов, недостаток которых, как и отсутствие единства исследователей по поводу морфологической близости рассматриваемых находок Африки и Азии, мешает сделать дефинитивное заключение.
Если не все ясно относительно морфологической общности потомков первых мигрантов, то, во всяком случае, нельзя отрицать их хронологическое единство.
Тогда встает вопрос о том, случайна ли эта близость во времени? Здесь необходимо обратиться к южноафриканским находкам SK 847 и Stw 53, оценка таксономического статуса которых до сих пор вызывает разногласия и колеблется между Н. habilis и Н. erectus/ergaster. Дискуссия по поводу эволюционного статуса SK 847 имеет особенно долгую историю. Первоначально известный исследователь австралопитековых Дж. Робинсон включал этот экземпляр в вид Homo erectus. Позже находка стала фигурировать как Homo ergaster (Johanson & Blake, 1996). Но Б. Вуд поместил ее в вид Homo habilis причем даже Н. habilis sensu stricto (Wood,1992). К этой оценке присоединился Ф. Грайн (Grine, 2000), обративший внимание на то, что сходство с Homo ergaster/erectus практически сводится к степени развития надглазничного торуса. Такой же таксономический диагноз Ф.Грайн дал и в отношении экземпляра Stw 53, подчеркнув при этом его большое сходство с SK 847. Заметим, что древность этих экземпляров — 1,8 млн. лет — совпадает с датировками находок в Дманиси (1,77 млн. лет). К сожалению, упомянутые выше дискуссионные экземпляры фрагментарны, однако все же их в какой-то мере можно использовать для сопоставлений. На рис. 4 изображен череп (фрагмент) SK 847 (Сварткранс) в сравнении с черепом D2700 (Дманиси). Бросается в глаза ряд общих черт: наклон контура части черепной крышки, резко выступающая латерально скуловая кость, глубокая вырезка скулового отростка верхнечелюстной кости, обширная и глубокая fossa canina, резкий перегиб в области скуло-челюстно-го шва. Различие в основном сводится к несколько сильнее развитому надглазничному торусу у SK 847. Спорный таксономический диагноз обеих форм объясняется их промежуточным эволюционным статусом, который может найти отражение в выделении предложенного выше отдельного подвида. Этот промежуточный тип раннего человека сформировался в Африке на позднем этапе периода «колыбели». О его африканском происхождении мы можем с большей определенностью судить в настоящее время на основании данных международной экспедиции, работавшей в южной Эфиопии (Консо, Konso-Gardula) и опубликовавшей в японском журнале Anthropological science (август 2007) большую статью (Suwa et al., 2007). Авторы рассматривают 8 фрагментов черепов (включая зубы) и подчеркивают, что изученный материал, отнесенный к раннему Н. erectus, обнаруживает мозаику примитивных и прогрессивных особенностей, причем может рассматриваться в рамках единой морфологической последовательности в течение сотен тысяч лет (1,65-1,0 млн. лет). Выдвигается гипотеза, согласно которой популяции африканских Н. erectus образуют вариабельную, но непрерывную преемственность, которая явилась источником многократных генных потоков на Евразийский континент («…multiple events of gene flow to the Eurasian continent» (Suwa et al., 2007, p. 133). Авторы рассматриваемой работы подчеркивают, что находка в Консо принадлежит к таксону Н. erectus в широком смысле и при этом фактически уравнивают термины Н. ergaster и Н. erectus … «earliest Н. erectus (or Н. ergaster)» (p. 134). Правда, H. erectus в строгом понимании считается более специализированной формой, связанной с Азией.
Для поставленной в моей работе задачи наиболее важен вывод о физическом типе первых мигрантов из Африки, который сформировался в период становления нового таксона на базе местного, африканского, что подтверждается либо очень малой хронологической трансгрессией

Сравнение черепов SK 847 (справа) и D 2700 (слева) (ссылки на литературные источники см. выше.
Н. habilis sensu lato и Н. erectus sensu lato, либо полным ее отсутствием (op. cit., р. 135).
Отметим, что сравниваемые находки ископаемых гоминид (SK847, Дманиси) связаны с миграциями, которые относятся к одному периоду времени и могут быть рассмотрены как представители самой ранней внутриафриканской и экстра-африканской дисперсии рода Homo. Таким образом, имеющийся антропологический материал в какой-то мере дает возможность ответить на вопрос «кто?» (т. е какие гоминиды) мог совершить «исход» гоминид из Африки. Его осуществили представители древнего подвида еще не сформировавшегося окончательно вида Homo erectus (в данной работе он предварительно назван Н. habilis pre-erectus). Я предпочитаю выделить самостоятельный подвид, а не просто констатировать промежуточный характер формы, как бы выхваченной из непрерывной последовательности, не только в силу наличия определенного морфологического комплекса, но в значительной степени в связи с фактами первых миграций, показавших, что этот комплекс сформировался и показал себя достаточно дееспособным для того, чтобы быть специфической поведенческой характеристикой отдельного таксона.
В данной работе (см. выше) был отмечен важный факт совпадения датировок древнейших остатков самых ранних эректусов в Африке и за ее пределами (SK 847, Stw 53, Дманиси, Ява). Ссылаясь на эти находки, объединяемые в особый таксон, вероятно осуществивший первый выход гоминид за границы Африки, мы везде сталкиваемся с датой 1,8 млн. лет, что позволяет предположительно ответить на вопрос о времени т. н. «исхода» человека с территории прародины. Эту цифру мы встречаем и в работах ряда исследователей (Бар-Йозеф, 1997, Antón et al., 2000). Учитывая время, необходимое для достижения новых земель, следует несколько удревнить предполагаемые сроки этого события — до 1,9 млн. лет, тем более, что не исключены новые открытия свидетельств пребывания человека в Азии, раньше предполагаемой на данный момент даты. Мнения исследователей, считающих, что «исход» имел место существенно раньше, чем думали прежние авторы, а именно — вскоре после хронологического рубежа 2 млн. лет — «soon after 2 т. у. а.» (Swisher et al., 1994; Mithen & Reed, 2002) или около 2 млн. лет назад (Деревянко, 2006) — оправдываются. Конечно, нужно учесть, что дискуссия о времени первого проникновения человека в Евразию не закончена, и существует довольно значительный разброс мнений на этот счет. Так X. А. Амирханов (1997) датирует это событие периодом времени 1,65-1,35 млн. лет, С. Гембл (по работе М. Волпоффа, 1997) называет дату 1,2 млн. лет до современности, С. Митен и М. Рид (Mithen & Reed, 2002) занимают крайнюю позицию, удревняя возможную датировку «исхода» до 2 млн. лет. Как во многих других подобных вопросах, сложились два альтернативных подхода исследователей к определению важной даты в эволюции человека — «короткая хронология» и «долгая хронология». На этот раз сумма аргументов разных областей науки скорее свидетельствует в пользу «долгой хронологии». Формирование нового подвида — Н. habilis pre-erectus, — его морфофизиологическая и поведенческая подготовленность к дальним переходам, сравнительно легкая доступность (в те времена) азиатского берега, видимого со стороны Эфиопского нагорья, сходство природных условий обоих берегов должны были «спровоцировать» проникновение гоминин в Азию в очень давнее время — примерно 2 млн. лет назад плюс-минус 0,2 млн. лет. С точки зрения данных современной науки другие варианты были бы странными и мало вероятными. Уточнение датировок и новые подходы к изучению ископаемых материалов позволяют внести коррективы в исследование древнейших этапов эволюции человека (Manzi, 2000). Комплексный подход к изучению таких сложных событий, как «исход» гоминид из Африки, выявил наличие определенной корреляции между физическим типом и миграционной активностью древнейших Homo и показал связь между формированием Homo erectus и резким подъемом подвижности популяций человека.
Далее речь пойдет о том, где, в какой точке границ Африканского континента, произошел первый выход древнего человека в Евразию.
Для начала уточним, в какой части света вне Африки впервые мог появиться человек — в Европе или в Азии. Третьего варианта, как говорится, не дано.
Этот вопрос неоднократно обсуждался специалистами, опубликовавшими ряд статей, в которых точки зрения авторов наглядно иллюстрируются картами предполагаемых путей расселения древнейших гоминин.
Посмотрим на карту (стр. 42), отражающую позицию разных специалистов по поводу движения популяций древнейшего человека после «исхода» из Африки. Наиболее подробно и убедительно выглядит картина выхода первых человеческих популяций в Азию, однако она не исключает и европейский путь в Евразию. Датировка стоянки Айн Ханех в северном Алжире (до 1,9 млн. лет) показывает что человек на эволюционном уровне раннего эректуса (или пре-эректуса) достиг территории нынешнего Магриба и находился уже у самого Гибралтарского пролива, «на пороге» Европы, в плио-плейстоценовое время, в период активных внуриафриканских миграций. Если бы в ту эпоху какие-то группы пре-эректусов преодолели зону Гибралтарского пролива, двигаясь на север, мы могли бы с достаточно высокой долей вероятности сказать, что второй частью света, которую посетил первый человек, была Европа. Возможность такой миграции, казалось бы, подтверждается наличием на юге Европы африканских животных, в частности, — обезьян Theropithecus oswaldi, — проникших из Африки в период существования «Гибралтарского моста», т. е. участка суши, соединявшего два континента в верхнем миоцене. Есть сведения, что такой «мост» сохранялся довольно долго, даже после 2,5 млн. лет до современности, но гоминины подошли к этому межконтинентальному рубежу, «опоздав» на несколько сотен тысячелетий. Впрочем, не только это «опоздание» помешало им распространить свою дисперсию на территорию Европы. Анализ расселения человека показывает, что древнейшие миграции этого рода имели в основном широтное направление и были ограничены тропическим и субтропическим поясами — от 35° с.ш. до 30° ю.ш., т. е.практически между Айн Ханех и Стеркфонтейн (Dennell, 2003). Это вполне понятно, учитывая уровень развития материальной культуры первых людей, который был еще недостаточен для адаптации к новым, более суровым и изменяющимся условиям Европы того времени. Были и другие факторы, лимитирующие неограниченную дисперсию групп гоминин, в частности уменьшение плотности населения при быстром распространении: это могло создать проблемы при подборе брачных партнеров (Dennell, op. cit). Во всяком случае, движение в северном направлении за пределы Африки в Европу не могло стать приоритетным для первых Homo. Впоследствии «Гибралтарский мост» распался, причем возник остров, соединившийся с Европой только в плейстоцене (Тернер, 1997). Прошел еще миллион лет прежде чем новая волна мигрантов Н. erectus/ergaster (или H.heidelbergensis) воспользовалась таким путем, но это уже были довольно поздние гоминины, не связанные с интересующим нас в данный момент периодом «исхода» из Африки. Правда, поиски остатков первых людей позволяют предположить, что человек, возможно, проник на Пиренейский полуостров раньше, чем принято считать (достоверная древнейшая дата — 0,9 млн. лет назад): испанские антропологи объявили о находке в районе Атапуэрка фрагмента черепа, возраст которого равен 1,3 млн. лет. Но все же, и эта новая находка значительно «моложе» первого «исхода» человека из Африки. Был еще один возможный путь проникновения человека в Европу из Африки — с территории современного Туниса — в Сицилию и далее — на Апеннинский полуостров по цепочке островов. Кстати, в Сицилии найдены примитивные орудия типа Mode 1.Однако, по датировке они опять-таки не увязываются с периодом первоначального «исхода» из Африки. Гипотеза более ранних посещений человеком Европы сталкивается с серьезными трудностями и не является общепринятой. Но если даже принять предельные называемые некоторыми исследователями спорные даты (1,6 млн. лет), то все равно получается, что человек проник в Европу существенно позже, чем в Азию. Древнейших по глобальному масштабу свидетельств пребывания человека в Европе пока не обнаружено.
На упомянутой карте (рис. 5) четко обозначены два возможных пути перехода древнейших гоминин из Африки в Азию: северный путь по долине Нила до Синайского п-ова и восточной части Средиземного моря и южный — через Баб-эль-Мандебский пролив. При обширной дисперсии древнейшие гоминины могли воспользоваться обоими путями, но все же, учитывая основную идею данной работы, будет полезно попытаться представить себе, на основании имеющихся данных, какой из этих двух вариантов могли избрать древнейшие люди в первую очередь.
Миграция по долине Нила к Средиземному морю имела некоторые преимущества. Прежде всего она, по утверждению исследователей, была для древнейших людей «более безопасной» (Бар-Йозеф, 1997, стр.47), (правда автор цитируемой работы в данном случае подробно не обосновал такой вывод). Против этой гипотезы можно было бы выдвинуть рассмотренное выше соображение, касающееся приоритетного характера широтного направления движений древних гоминин и трудностей продвижения на север. Однако, этот аргумент, имеющий существенное значение в отношении перехода в Европу, теряет силу при анализе миграций в пределах Африки, причем в основном южнее 30° с.ш., в очень мало меняющихся природных условиях. Здесь следует задуматься над вопросом, что могло привлекать мигрантов на северном пути. Прежде всего здесь может идти речь о наличии постоянных ресурсов пресной речной воды. Однако, тогда непонятно, зачем древним Homo понадобилось уходить из района озера Туркана, а не наоборот — стремиться к берегам этого озера. Судя по карте, опубликованной в работе Р.Деннелла (Dennell, 2003, р.427) (рис. 5), большая часть территории Восточной Африки, прилегающая к Красному морю и территории современной Эфиопии в целом в позднем плиоцене представляли собой саванны с отдельными участками вечнозеленых лесов, что свидетельствует о достаточном количестве воды в рассматриваемом регионе, находящемся в непосредственной близости от бассейна Нила. Значит, были другие факторы, обусловившие перемещение популяций гоминин. При этом движение на север, казалось бы, не давало людям каких-либо новых жизненно важных выгод. Если в Египте несколько тысячелетий до современности заселение долины Нила стимулировалось наличием плодородных почв, обусловивших развитие земледелия, то два миллиона лет назад этот фактор, естественно, не имел никакого значения. Вероятно, описываемые события связаны главным образом с пищевой стратегией. Речь идет о каких-то группах самых ранних представителей, рода Homo о том динамичном подвиде, который в этой работе мы называем Н. habilis pre-erectus. Представители этого таксона не были еще охотниками, — мы определяем их как активных («конфронтационных») скэвинджеров. Необходимо учесть, что этот тип пищевой стратегии в описываемый период претерпевал существенные изменения в связи с массовым вымиранием крупных хищников в период от 2 млн. до 1 млн. лет назад и расширением ниши гоминин в отношении пищевого поведения (Lewis, 1997). Зависимость людей от хищников уменьшилась, но, в то же время, гоминины претерпели определенный ущерб в связи с исчезновением «поставщиков» больших количеств калорийной пищи, важную в качественном отношении которой часть составлял костный мозг. Говоря о пищевых стратегиях Homo, нельзя забывать, что этот таксон никогда не был узко специализированным: диэта этих гоминин всегда была достаточно разнообразной и включала различные виды пищи, в частности такие, которые доставлялись собирательством. В числе последних важную роль, вероятно, играли моллюски, в основном морские, обитающие в прибрежной полосе и представлявшие легкую добычу для собирателей, которые имели таким образом постоянный источник «ценной»

Распределение растительности в северном полушарии в позднем плиоцене — раннем голоцене. R. Dennell: «Dispersal and colonization, long and short chronologies: how continuous is the Early Pleistocene record for hominids outside East Africa?». Journal of Human Evolution. 2003. N 45. P. 421–440.
высоко калорийной пищи, необходимой для существ с развитым мозгом. Но чтобы овладеть этими практически неограниченными пищевыми ресурсами нужно было выйти к морю. И, разумеется, для первых Homo легче всего и раньше всего должны были стать доступными берега Красного моря и Аденского залива. Путь до Средиземного моря был гораздо более далеким. Конечно, древнейшие люди не планировали заранее «экспедиции» к морю. Просто морской берег в этом регионе Африки лежал на пути их первых, даже самых близких миграций, так что представители ранних Homo уже вскоре после формирования древнейших вариантов вида могли приобрести комплекс «береговой адаптации» и стать собирателями прибрежных моллюсков. В эпоху плио-плейстоцена и позже берег Красного моря со стороны Эфиопии был легко доступен: на месте нынешней пустыни Данакиль простиралась полоса саванн, и древние гоминины хорошо освоили приморскую область. Это, в частности, доказывает находка в Эритрее черепа Homo erectus/ergaster, получившего по каталогу шифр Данакиль UA31. Древность находки — более 1 млн. лет (Abbate et al., 1998). Череп имеет сильно развитый надглазничный торус, сравнимый по измерительным данным с аналогичным параметром Сангиран 17, однако отличается сходством по ряду признаков с Н. ergaster (Manzi, 2000).
Имеющиеся данные антропологии и археологии позволяют сделать вывод, что древние гоминины уже начиная с плио-плейстоценового времени были «у ворот Азии» на юге побережья Красного моря, т. е. в районе нынешнего Баб-эль-Мандебского пролива. Этот факт, разумеется, привлекает внимание исследователей, занимающихся проблемой древних миграций человека. Так, X. А. Амирханов пишет, что «…вновь выносится на обсуждение вопрос о кратком пути расселения людей в Азию через территорию современного Баб-эль-Мандебского пролива, как дополнение или альтернативу длинного пути через Эфиопское нагорье и долину Нила» (Амирханов, 1997 стр. 66). В работе 1991 г. этот же автор дает важную для интересующей нас проблемы «исхода» гоминин из Африки информацию о том, что на юге Аравийского полуострова в раннем палеолите некоторые районы были благоприятными для обитания, причем «располагались в пределах видимости от Афарского рифта в Эфиопии, от которой они отделялись только узким и мелководным проливом Баб-эль-Мандеб» (Амирханов, 1991, цитируется по работе Бар-Йозеф, 1997, стр. 50). То обстоятельство, что азиатский берег был видим с территории африканской «колыбели» человечества, могло играть существенную роль в качестве психологического стимула перехода в Азию, тем более, что в природном отношении территория юга Аравийского полуострова практически должна была восприниматься как продолжение столь знакомой Африки. Естественно предположить, что именно в районе Баб-эль-Мандебского пролива произошло событие, которое служит центральной темой данной главы — первый «исход» древнейших людей из Африки. Такую же точку зрения высказывают сейчас некоторые зарубежные антропологи и археологи. Р. Деннелл пишет, что «барьер Красного моря между Африкой и Аравийским полуостровом, вероятно, был гораздо менее эффективным, чем теперь, особенно на юге, у пролива Баб-эль-Мандеб (Tchernov, 1992), который мог быть важным местом перехода (crossing point) между Африкой и Азией» (Dennell, 2003), и у гоминин не было причин для прекращения дальнейшей дисперсии в такие же как в Африке саванны Азии. Согласно мнению другого английского археолога А. Тернера «можно предположить, что в позднем плиоцене происходили миграции между восточной Африкой и южным Левантом через Баб-эль-Мандебский (африканско-арабский) мост, существовавший на самом юге Красного моря, несмотря на то, что в это время уже происходило его расширение (Tchernov, 1992)» (Тернер, 1997, стр. 14). Замечание относительно расширения Красного моря дает дополнительную возможность представить себе географию района Баб-эль-Мандебского пролива в плио-плейстоценовое время: пролив был очень узким и действительно, как считает Р. Деннелл, не представлял серьезного препятствия. Кроме того, в силу понижения уровня моря на 150 м после временного интервала верхний плиоцен— ранний плейстоцен (Амирханов, 1997) глубина пролива стала небольшой. Надо также учесть возможность существования отдельных участков суши в его пределах. В настоящее время, например, в средней части Баб-эль-Мандебского пролива существует остров (Перим).
В итоге складывается представление, основанное на данных палеогеографии, палеоэкологии и палеоантропологии, что путь древнейших гоминин из Африки в Азию через область Баб-эль-Мандебского пролива был приоритетным во времена «исхода» и, более того, был практически единственным для самой ранней миграции, которая, по нашему мнению, осуществлялась еще до формирования «типовых» Homo erectus представителями подвида
Н. habilis pre-erectus, проявившего в те времена высокий уровень миграционной активности, судя по находкам в Дманиси и Южной Африке (SK 847, Stw 53). Английские исследователи С.Митен и М. Рид (Mithen & Reed, 2003) приводят на стр. 434 своей работы очень выразительную карту предполагаемого «исхода» древнейшего человека (рис. 2). Обратим внимание на соотношение северного и южного путей этой ранней миграции гоминин. На карте не изображен отрезок пути из Эфиопии до Синайского полуострова — здесь авторы проявляют определенную научную осторожность. Зато южный путь обозначен максимально четко и однозначно: он проходит через область Баб-эль-Мандебского пролива, идет через Аравийский полуостров и уводит прямо на восток, достигая территорий Китая и Индонезии. Северный участок пути поворачивает к северу, через Кавказ (Дманиси) и продолжается в сторону Европы, нигде не уклоняясь на восток. Основная идея схемы сводится к тому, что «исход» через Баб-эль Мандебский пролив являлся единственным для описываемого времени направлением на восток и был стартовым моментом грандиозной миграции через всю территорию Азии с образованием «восточного очага» антропогенеза. В рассматриваемой работе С. Митен и М. Рид на стр. 456 прямо сказано, что компьютерное моделирование по программе STEPPINGOUT приводит к выводу, что «афарский путь» из Африки, т. е. через Баб-эль-Мандебский пролив, имел большее значение для гоминид, чем расселение через долину Нила. Авторы не проводят линию, ведущую на восток по южному берегу Аравийского полуострова, заменяя ее резким поворотом на север и далее — в обход Персидского залива. Это наводит на мысль, что этим же путем теоретически можно было бы сравнительно легко достигнуть Ближнего Востока. Тогда «исход» через Баб-эль-Мандебский пролив остался бы вообще единственным предполагаемым вариантом проникновения ранних Homo в Азию. Однако, С. Митен и М. Рид все же предпочитают сохранить идею северного пути в Азию наряду с южным, причем, возможно, они правы. Но в данной работе для нас важен вопрос о первом во времени эпизоде «исхода», и в этом отношении Баб-эль-Мандебский путь, как говорится, «вне конкуренции». Во всяком случае, таким нам представляется одно из наиболее вероятных решений рассматриваемого вопроса.
Работа исследователей над рассматриваемой проблемой «исхода» гоминин из Африки продолжается и, надо сказать, приносит новые подтверждения гипотезы «афарского» (т. е. южного) пути первоначального расселения гоминин из Африки в Евразию. В 2007 году опубликована статья группы английских ученых (Hughes et all., 2007), осуществивших детальный анализ природных условий в эпоху плио-плейстоцена в разных регионах Земного шара, в том числе — в области Эфиопского нагорья и Баб-эль-Мандебского пролива. Применен усовершенствованный метод компьютерного моделирования. Исследование показало, что к северу от пролива, начиная примерно со среднего течения Нила, вплоть до Синайского перешейка, простиралась обширная пустыня, очевидно совершенно непроходимая для ранних Homo. В то же время на юге побережья Красного моря была саванна а непосредственно в области Баб-эль-Мандебского пролива оставались еще участки тропического леса. Можно предположить, что стволы деревьев, падавшие в воду могли помочь древним людям переправиться через пролив, который, сильно обмелел в те времена. Новое исследование показало, что уровень моря был тогда на 120 м ниже современного, который, кстати, тоже не превосходит 200 м. В общем, все больше данных свидетельствует в пользу «афарского» пути первого «исхода» гоминин из Африки. Поданным английских исследователей группы древнейших людей, переселившихся в Азию, очевидно, двигались «в обход» Аравийского полуострова, после чего для них открылось направление на Ближний Восток и на Кавказ.
Нам могут возразить, что на Ближнем Востоке имеются свидетельства очень раннего проникновения человека, например стоянка Эрк-эль-Ахмар (1,9 млн. лет) и даже еще более древние находки (Тернер, 1997), что может в территориальном отношении быть связано с северными регионами Африки. Несомненно, такие факты могут рассматриваться в едином контексте с североафриканской дисперсией Н. erectus, оставившей, в частности, стоянку Айн Ханех. Но ведь это может служить доказательством возможности «исхода» из Африки на севере через область Суэцкого перешейка, но ничего не говорит о невозможности более раннего «исхода» через Баб-эль-Мандебский пролив. Вопрос здесь сводится к дилемме, касающейся движения гоминин до Синайского полуострова вдоль западного или вдоль восточного берега Красного моря. Последний вариант означал бы «привязанность» первой миграции к области Баб-эль-Мандебского пролива и признание за этим участком Восточной Африки роли единой точки «исхода» первых ранних Homo из «колыбели» этого рода.
Разумеется, говоря об «исходе» как одной неповторимой точке в пространстве-времени, мы имеем в виду только самое раннее событие у истоков человечества. Исследователи не раз высказывали предположение о множественном характере феномена «исхода». Об этом писал Р. Деннелл (Dennell, 2003), а итальянский археолог К. Перетто (Peretto, 2000) конкретно назвал две предполагаемые «волны» расселения из Африки: первую, раннюю, принесшую индустрию Mode 1 и вторую, среднеплейстоценовую, связанную с носителями ашельской техники Mode 2. Вообще, с тех пор, как человек приобрел морфофизиологический и поведенческий комплекс древнейших Homo, обеспечивший ему возможность экспансии в широком масштабе, такие «прорывы» за пределы Африки должны были стать скорее обычными эпизодами, а не уникальными случаями. Одновременно шел процесс все более углубленного познания и освоения Африканского континента. Естественно, нельзя думать, что при миграциях, даже самых значительных, прародину покидал целый таксон. Часть мигрирующей популяции, как правило, остается на месте (Бар-Йозеф, 1997). Группы Homo habilis pre-erectus, сформировавшегося в «колыбели» на территории нынешней южной Эфиопии и Кении, продолжали эволюционировать в прежнем направлении, о чем свидетельствуют такие палеоантрпологические находки, как скелет из Нариокотоме (Кения, 1,6 млн. лет), череп KNM-ER 3733 (Кения, 1,5 млн. лет), фрагменты черепа из Консо Гардула (Эфиопия, 1,4 млн. лет). Эти материалы свидетельствуют о преемственности эволюционных стадий Н. erectus на территории прародины последнего и об окончательном становлении этого вида, который некоторые исследователи считают «первым настоящим человеком».
Этапы становления Homo erectus/ergaster хорошо прослеживаются на краниологических материалах, относящихся к периоду 2+ — 1,5 млн. лет до современности. На это время приходится целый ряд находок «промежуточного» характера, вызывающих постоянные споры по поводу их систематического положения (рис. 6 a-d). Древнейшим звеном мы в данном случае будем считать KNM-ER 1470, который сейчас относят к Homo rudolfensis, т. е. к эволюционному уровню хабилиса, хотя в его морфологических особенностях исследователи всегда отмечали присутствие черт Homo erectus (рис. 6 а). Здесь перед нами — фактически уже знакомая проблема таксономического диагноза древнейших Homo, на деле являющихся ранней формой Homo erectus, которую мы определили как Homo habilis pre-erectus. — предшественника H. erectus. Ее полный морфологический комплекс проявился в черепной серии из Дманиси. Поэтому нет необходимости возводить № 1470 в ранг отдельного вида, что было сделано в то время, когда еще не было ключевого материала для выделения раннего подвида Homo erectus. Этот экземпляр, по нашему мнению, должен быть включен вместе с находками в Дманиси и южноафриканскими черепами SK 847 и Stw 53 в подвид Homo habilis pre-erectus, «типовым» экземпляром которого является череп D2700 из Дманиси (рис. 6 b). Часть древней популяции ранних Homo продолжала эволюционировать на территории Кении и Эфиопии, где примерно 1,6 млн. лет назад выделился вариант KNM-WT 15000 — Нариокотоме III (рис. 6 с), отличающийся определенным своеобразием, особенно в том, что касается посткраниального скелета (большая длина тела). Иногда эту находку прямо относят к Homo ergaster, но достаточно посмотреть на череп KNM-WT 15000 в латеральной норме, чтобы убедиться в его очевидном сходстве с Homo hablis pre-erectus. Это сходство проявляется также в морфологии зубов: уникальная форма верхних резцов, коронка которых сильно расширяется в сторону режущего края отмечается на черепах Нариокотоме и Дманиси D2700; очень большая ширина тригонида по отношению к талониду характерна для первых нижних моляров Нариокотоме и D2735. Речь здесь идет о редких признаках, что наводит на мысль об общих генетических корнях палеодема Дманиси и ранних африканских Homo. Эволюционный ряд краниологических типов от истоков рода Homo до дефинитивных форм Homo erectus/ drgaster завершает «типовой» для Homo ergaster череп KNM-ER 3733 из Кении (рис. 6 d). По объему мозговой коробки (848 см3) этот экземпляр мало отличается от Нариокотоме (880 см3), но свод черепа у него выше, а чешуя лобной кости, отделенная от надглазничного валика глубокой канавкой, гораздо менее наклонна, чем на черепах Homo habilis pre-erectus. Граница между этими таксонами определяется не только по данному признаку: в Рассмотренной работе о черепах из Дманиси (Rightmi-er et al., 2006) авторы провели сравнительное исследование черепов ранних гоминид по параметрам строения нёба, причем был построен бивариантный график, на координатах которого отложены глубина нёба и нёбный индекс (р. 132). Точки, обозначающие черепа KNM-ER 3733 и D2700 расположились на поле графика на очень значительном расстоянии друг от друга. Перед нами наглядная картина становления нового таксона африканских Homo, который многие исследователи сейчас определяют как вид Homo ergaster, но автор данной статьи в интересах последовательности своей позиции в систематике гоминин предпочитает рассматривать термин «ergaster» скорее как подвидовой, входящий в триномен Homo erectus ergaster. Это наименование, кстати, совместимо с позицией тех антропологов, которые предпочитают не противопоставлять эволюционный уровень форм ergaster и erectus, считая биномен Homo erectus, универсальным для африканских и азиатских гоминин, относящихся к стадии «первых настоящих людей». В цитируемой статье, посвященной человеку из Дманиси, это проявляется
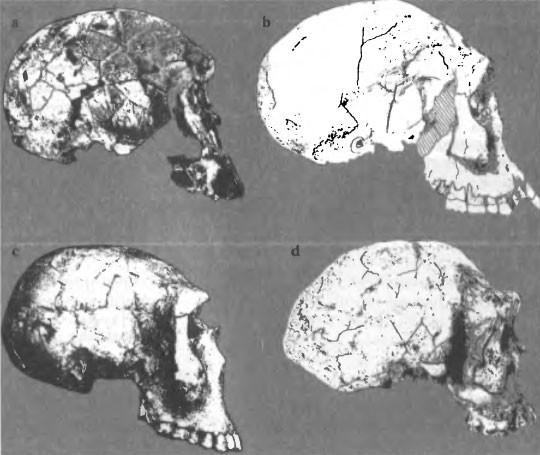
Эволюция черепа Homo erectus / ergaster от древнейших форм (KNM-ER 1470) (а) через «типовой» вариант» Н. habilis pre-erectus (b) (Дманиси D2700), промежуточный тип Нариокотоме III (с) — до «классического» Homo erectus ergaster KNM-ER 3733 (d)
вполне отчетливо: на стр. 133 читаем «…African erectus (=ergaster)». Подвидовой уровень различий между африканскими и азиатскими Н. erectus авторы связывают с большой, по их мнению, скоростью первоначального расселения ранних Homo после «исхода» из Африки и недостатком времени для возникновения существенных таксономических различий («…there would have been little time for the diverging groups to develop distinctive characters» («…дивергирующие группы имели, вероятно, мало времени чтобы выработать отличительные признаки». (op. cit. р. 139). Выше приводились рассуждения, согласно которым незначительная интенсивность процессов дивергенции и видообразования у человека может быть объяснена на основании специфических особенностей рода Homo. Все же пока сходство африканских и азиатских эректусов на подвидовом уровне не будет твердо доказано (а у меня остаются некоторые сомнения на этот счет), «не следует недооценивать определенных различий между сравниваемыми формами» (Васильев, 1999).
В итоге, генеалогическое древо рода Homo может выглядеть, по нашим представлениям, следующим образом (рис. 7). Род Homo выделяется из среды австралопитековых как Homo habilis sensu lato около 2,5 млн. лет назад и через 200–300 тыс. лет происходит разделение этого таксона на две ветви: Homo habilis habilis et Homo habilis pre-erectus. Последний широко расселяется го территории Африки и осуществляет «исход» — на земли Евразии, сравнительно быстро (?) достигая Юго-Восточной и Восточной Азии, сохранив при этом видовую (erectus) и, отчасти, подвидовую модель (mojokertensis ~ pre-erectus). Далее происходит дивергенция подвида Н. erectus erectus и его восточноазиатского ответвления — Н. erectus pekinensis. Лантьянслого эректуса, очевидно, можно отнести к Н. habilis pre-erectus (mojokerlensis),
В Африке анагенегическая эволюция приводит к становлению Н. erectus ergaster, который может рассматриваться как «opposite number» азиатского Homo erectus erectus. От него происходят Н. erectus mauretanicus и Н. antecessor (=ранний H. heidelbergensis).
Возникновение нового таксона, достигшего максимально совершенного уровня адаптации в данных условиях, часто связано со «взрывами» миграционной активности. Во всяком случае, на протяжении всей линии эволюции рода Homo эта закономерность проявляется на самых различных уровнях.
Существует мнение, согласно которому граница Африканского континента в палеолите пересекалась не только несколько раз в направлении «out-of Africa», но также были возможны миграции противоположного направления — «back-to-Africa» (Wood & Turner, 1995). Аргументация этой точки зрения строится на фактах наличия, по мнению некоторых антропологов, морфологических черт Н. erectus sensu stricto на черепах, по времени относящихся к периоду 0,9–1,2 млн. лет до современности, найденных на территориях Европы (Чепрано) и восточной Африки (ОН 9). Б. Вуд и А. Тернер предположили, что древнейшие Homo вышли из Африки в позднем плиоцене, а Н. erectus эволюционировал в Азии, но потом вернулся в Африку (Wood & Turner, 1995). Антропологи и археологи последующих лет в основном положительно восприняли идею возможности возникновения Н. erectus в Азии (Hyodo et al., 2002, Деревянко, 2006). Исследователи справедливо замечают при этом, что признание этой гипотезы в сильной степени зависит от даты первоначального появления Homo в Азии, а, надо сказать, что в этом вопросе существует довольно широкий спектр мнений. Если исходить из наиболее распространенной сейчас датировки (Swisher et al., 1994), то «сценарий» Б. Вуда и А. Тернера представляется вполне правдоподобным: человек пришел в Индонезию на ранней стадии еще не сформировавшегося вида Н. erectus и эволюционировал как Н. erectus erectus, в то время как в Африке развился вариант Н. erectus ergaster. Однако, гипотеза «back-to-Africa» пока не получила широкого распространения среди специалистов. Представляется мало понятным «поворот на 180°» самой дальней

Предлагаемая схема систематики и эволюции рода Homo.
миграции рода Homo, популяции которого должны были хорошо адаптироваться за несколько сотен тысяч лет в Юго-Восточной и Восточной Азии, так что, во всяком случае на первый взгляд, у них не могло быть никакого стимула для ухода на запад. Но при этом надо иметь в виду, что заранее запланированного направления движения у древних гоминин не было, и на огромном протяжении пути из Африки в Юго-Восточную Азию вектор дисперсии мог меняться. Упоминавшиеся выше английские антропологи С. Митен и М. Рид, разрабатывая принципы компьютерного моделирования миграций древнего человека, исходили из постулата, что при отсутствии предопределенного маршрута расселения через Евразию «…hominids can just as easily spread back into Africa» («…гоминиды могли столь же легко распространяться назад в Африку» (Mithen & Reed, 2002, p. 437). Признавая справедливость этого замечания, не следует все же забывать, что мигранты, хотя бы в течение многих поколений, все же достигли Индонезии, а значит в среднем преимущественное направление на восток очень долгое время довольно устойчиво сохранялось, о причинах чего остается лишь строить гипотезы. Факты пока свидетельствуют о длительном предпочтении восточного направления в период рассматриваемой миграции и не дают основания для альтернативного предположения о сколько-нибудь стабильном западном векторе расселения. Можно, конечно, представить, что формирование вида Homo erectus erectus вызвало новый демографический «взрыв» и, соответственно, — новую волну экспансии на территории Азии в период 1,2–0,9 млн. лет, с дисперсией во все стороны, причем такая гипотеза легче доказуема в отношении движения на север, но для утверждения реальности западного вектора пока нет достаточного основания: новый вид возникает в Восточной или Юго-Восточной Азии, появляется в Африке и Европе, а на среднем участке «обратного» пути через всю Азию антропологических свидетельств этих событий не найдено. Те же сомнения не исключены и в отношении древнейшей миграции на восток, но в этом случае они устраняются археологическими данными по Индии и Пакистану. Гипотеза «out-of-Africa» на данный момент выглядит более убедительно, чем вариант «back-to-Africa», хотя последний не исключается и показывает возможность многократного пересечения границ Африки уже на стадии пре-эректуса после первого древнейшего «исхода».
1. «Исход» из Африки древнейших представителей рода Homo имел огромное значение для человечества и явился существенным этапом эволюции, т. к. положил начало глобальной дисперсии гоминин и связанной с ней морфофизиологической и поведенческой универсализацией адаптивного потенциала, выразившейся прежде всего в развитии мозга, расширением когнитивных возможностей и, соответственно, технологическом прогрессом.
2. Расселение древнейших Homo за пределы Африки стало возможным и необходимым по целому ряду причин, среди которых можно выделить: резкое изменение климата в период 2,5–2,0 млн. лет до современности (глобальный фактор), переход ранних Homo к новой пищевой стратегии (мясная пища, «конфронтационный скэвинджинг»), приобретение адаптации к открытым пространствам, включая бег на большие расстояния, соответствующие морфофизиологические изменения (увеличение длины ног, общих размеров тела, совершенствование терморегуляции), опыт, накопленный при освоении Африканского континента, рост численности популяции и расширение площади занимаемой отдельными группами.
3. Эволюция гоминин в Африке в период 2,4–1,8 млн. лет до современности привела к становлению нового вида человека — Homo erectus (sensu lato), который уже на самых ранних стадиях своего формирования приобрел более высокий, по сравнению с предками, уровень мобильности, давший возможность освоить (или, хотя бы, обойти) практически всю территорию Африки — от крайнего северо-запада (Алжир, Айн Ханех) до крайнего юга (Стеркфонтейн, Сварткранс), а затем — выйти за пределы континента (Дманиси, Моджокерто). Этот вариант раннего Homo, который известен нам по материалам со стоянки Дманиси (Грузия) и черепам из южной Африки (SK 847 и Stw 53), отличавшийся от сформировавшихся вариантов Homo erectus/ergaster рядом архаичных черт, сближающих его с Н. habilis sensu lato, может рассматриваться как самостоятельный таксон в пределах вида Homo erectus или Homo habilis. В первом случае его можно определить его как подвид — Homo erectus praevius, во втором — Homo habilis pre-erectus. Именно его представители стали «первооткрывателями» Евразии.
4. В науке в настоящее время существует довольно широкий спектр мнений по поводу датировки момента выхода рода Homo в Евразию — от 2 млн. лет до 1,2 млн. лет до современности. Автор данной статьи солидарен с позицией тех исследователей, которые оценивают древность рассматриваемого события формулировкой «сразу после 2 млн. лет назад» (Mithen & Reed, 2003), т. е. присоединяется к сторонникам «долгой хронологии». Если принять более позднюю дату, возникнут трудности с объяснением факта появления ранних Homo на Кавказе и на Яве в период 1,7–1,8 млн. лет до современности. «Долгая хронология» наилучшим образом согласуется с «возрастом» первых Homo, проявивших миграционный динамизм в панафриканском и экстра-африканском масштабе.
5. Дискуссионным является также вопрос о месте выхода древнейших людей за пределы Африки. Теоретически здесь возможны три ответа, подсказываемые самой географией континента: «исход» мог произойти в районах: 1. Гибралтарского пролива, 2. Суэцкого перешейка и 3. в области Баб-эль-Мандебского пролива на юге Красного моря. Первый вариант, скорее всего, должен быть исключен, по крайней мере для описываемого времени: эволюционный статус гоминин, включая низкий уровень материальной культуры, вряд ли позволил бы им переход на север, в Европу, связанный с резким изменением условий среды и, соответственно, со значительной перестройкой адаптации. Человек смог воспользоваться этим путем проникновения в Европу значительно позже. Из двух возможных точек «исхода» в Восточной Африке древнейшие Homo должны были в первую очередь предпочесть район Баб-эль Мандебского пролива, который был в то время уже и мельче, чем теперь и, что самое главное, находился вблизи «колыбели» первого человечества. Важно при этом, что азиатский берег был видим с территории Эфиопского нагорья и по своим природным условиям мало отличался от последнего. Палеоантропологические находки в этом регионе (Эритрея) свидетельствуют о том, что Homo erectus давно освоился в южной части побережья Красного моря и, вероятно, приобрел черты «береговой адаптации». «Исход» в этой части Африканского континента не должен был представить для древнейших жителей Эфиопского нагорья сколько-нибудь значительных трудностей, так что экспансия в восточном направлении стала первой и наиболее значительной экстра-африканской миграцией рода Homo, приведшей к его глубокому проникновению в Евразию. Позже несомненно имели место другие случаи выхода гоминин из Африки в разных ее регионах, т. е. правы также исследователи, считающие, что «исход» в пространственно-временном отношении носил множественный характер, но в данной работе ставилась задача выявить именно первое событие этого рода. В разнонаправленной первичной дисперсии человечества по разным причинам приоритетным направлением оказалось восточное, которое послужило основой разделения общего ствола эволюции на западную и восточную ветви.
Амирханов X. А. Палеолит Южной Аравии. М., 1991
Амирханов Х. А. Археологические данные проблеме аравийского пути первоначального рас: еления человечества. // Человек заселяет планету Земля. М., 1997. С. 65-77.
Бар-Йозеф О. Нижнепалеолитические стоянки Юго-Западной Азии — свидетельства расселения человека из Африки. // Человек заселяет планету Земля. М., 199. С. 41–65.
Бутовская M.Л. Социальная структура ранних гоминид и проблема адаптации к различным климатическим условиям в палеолите. // Человек заселяет планету Земля. М., 1997. С. 21–29.
Васильев С. В. Дифференциация плейстоценовых гоминид. М., 1999.
Волпофф М. Н. Из Африки. // Человек заселяет планету Земля. М., 1997. С. 29–41.
Деревянко А. П. Палеолит Китая: итоги и некоторые проблемы в изучении. Новосибирск. Издательство Института археологии и этнографии СО РАН. 2006.
Зубов А. А. Некоторые данные одонтологии к проблеме эволюции человека и его рас. // Проблема эволюции человека и его рас (коллективная монография). 1968. С. 5–124.
Зубов А. А. Тенденции эволюции человечества.// Расы и народы. 1982. Вып.12. С. 70–88.
Зубов А. А. Магистрализация и демагистрализация в ходе эволюционного процесса. // Вопросы антропологии. 1985. Вып. 75. С. 14–27.
Зубов А. А., Халдеева Н. И. Одонтология в современной антропологии. М., 1989. С. 1–218.
Зубов А. А. Палеоантропологическая родословная человека. М., 2004. С. 1–551.
Тернер А. Распространение ранних гоминид. // Человек заселяет планету Земля. М., 1997. С. 7–21.
Abbate Е., Albinelli A., Azzaroli A., Benvenuti М., Tesfamariani В., Bruni P., Cipriani N., Clarke R. J., Ficcarelli G., Macchiarelli R., Napoleone G., Papini М., Rook L., Sagri М., Tecle Т., Torre D., Villa I. A one-million-year-old Homo cranium from the Danakil (Afar) depression of Eritrea // Nature. 1998. Vol. 393. P. 458–460.
Antón S. C., Fachroel Aziz, Yahdi Zaim. Plio-Pleistocene Homo: Patterns and Determinants of Dispersal. // Humanity from African Naissance to Coming Millennia. Witwatersrand/Firenze. 2000. P. 91–102.
Antón S. C., Leonard W. R., Robertson M. L. An ecomorphological model of the initial hominid dispersal from Africa. // Journal of Human Evolution. N 43. P. 773–785.
Arcadi A. Clark. Species resilience in Pleistocene hominids that traveled far and ate widely: An analogy to the wolf-like canids. // Journal of Human Evolution. 2006. N. 51. P. 383–394.
Bailey S. E. & Lynch J.M. Diagnostic differences in mandibular P4 shape between Neandertals and anatomically modern humans. American Journal of Physical Anthropology. 2005. 126(3). P. 268–277.
Bailey S. E. Beyond shovel-shaped incisors: Neanderthal dental morphology in a comparative context. // Periodicum biologorums. 2006. Vol. 108. N 3. P. 253–267.
Boaz N. T. & F. Clark Howell A gracile hominid cranium from upper member G. of the Shungura formation, Ethiopia. // American Journal of Physical Anthropology. 1977. Vol. 46.N1. P. 93–108.
BrauerG. & Schultz M. The morphological affinities of the Plio-Pleistocene mandible from Dmanisi, Georgia. // Journal of human Evolution. 1996. 30. P. 445–481.
Dennell R. Dispersal and colonisation, long and short chronologies: how continuous is the Early Pleistocene record for hominids outside East Africa? // Journal of Human Evolution. 2003. N 45. P. 421–440.
Eswaran V., Harpending H., Rogers A. R. Genomics refutes an exclusively African origin of humans. // Journal of Human Evolution. 2005. 49. P. 1–18.
Grine F. E., Jungers W. L., Schultz J. Phenetic affinities among early Homo crania from East and South Africa. // Journal of Human Evolution/ 1996. N 30. P. 189–225.
Grine F. E. Implications of Morphological Diversity in Early Homo Crania from Eastern and Southern Africa. // Humanity from African Naissance to Coming Millennia. Witwatersrand/Firenze. 2000. P. 107–117.
Hughes J. K., Haywood A.,MHhen S. J., B. W. Sellwood P., Valdes P. J. Investigating early hominin dispersal patterns: developing a framework for climat data integration. // Journal of Human Evolution. Vol. 53. (2007). P. 465–474.
Hyodo М., Nakaya H., Urabe H., Saegusa H., Xue Shunrong, Yin Jiyun, Ji Xuepin Paleomagnetic dates of hominid remains from Yuanmou, China, and other Asian sites. // Journal of Human Evolution. 2002. N 43. P. 27–41.
Johanson D., Blake B. From Lucy to language. London, 1996.
Lewis M. E. Carnivoran paleoguilds: implications for hominid food procurement strategies. // Journal of Human Evolution. 1997. N.32. P. 257–288.
Manzi G. The earliest diffusion of the Genus Homo toward Asia and Europe: a brief overview // Humanity from African Naissance to Coming Millennia. Witwatersrand/Firenze. 2000. P. 117–125.
Martinón-Torres М., Bastir M., Bermúdez de Castro J. M., Gómez A., Sarmiento S., Muela A., Arsuaga J. L. Hominin lower second premolar morphology: evolutionary inferences through geometric morphometric analysis. // Journal of Human Evolution. 2006. 50. P. 523–533.
Merwe van der N., Thackeray J. F., Lee-Thorp J. A., Luyt J. The carbon isotope ecology and diet of Australopithecus africanus at Sterkfontein, South Africa. // Journal of Human Evolution. 2003. N 44. P. 581–597
Mithen S., Reed M. Stepping out: a computer simulation of hominid dispersal from Africa. // Journal of Human Evolution. 2003. N 43. P. 433–462.
Peretto С. The oldest human population in Europe: «long» or «short» chronologies. // Humanity from African Naissance to Coming Millennia. Witwatersrand/Firenze. 2000. P. 175–183.
Prat S., J.-Ph. Brugal, J.-J. Tiercelin, J.-A. Barrat, M. Bohn, A. Delagnes,
S. Harmand, K. Kimeu, M. Kibunjia, P.-J. Texier, H. Roche First occurrence of early Homo in the Nachukui formation (West Nurkana, Kenya) at 2.3–2.4 Myr. // Journal of Human Evolution. 2005. 49. P. 230–240.
Rightmire G. Ph., Lordkipanidze D., Vekua A. Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia. // Journal of Human Evolution. 2006. N 50. P. 115–141.
Stewart K. M. Early hominid utilization of fish and implications for seasonality and behaviour. // Journal of Human Evolution. 1994. 27. P. 229–245.
Suwa G., Asfaw B., Haile-Selassie Y., White Т., Katoh Sh., Woldegabriel G., Hart W. K., Nakaya H., Beyene Y. Early Pleistocene Homo erectus fossils from Konso, Southern Ethiopia // Anthropological Science. 2007. Vol. 115. P. 133–151.
Swisher C. C., Curtis G. H., Jacob Т., Getty A. G., Surpijo A., Widiasmoro Age of the earliest known hominids in Java, Indonesia // Science. 1994. Vol. 263. P. 1118–1121.
Tchernov E. (1992) Цитируется по работе: А. Тернер (1997) в сб. «Человек заселяет планету Земля» (М., 1997). Стр. 14.
Weidenreich F. The dentition of Sinanthropus pekinensis. Peking. 1937. Pal. Sinica N 5. P. 1–180.
Wood B. A. Origin and evolution of the genus Homo // Nature. Vol. 355. P. 783–790.
Wood B. & Turner A. Out of Africa and into Asia. // Nature. 1995. Vol. 378. P. 339–340.
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги