Глава V Новый Свет
В предыдущих главах речь шла об эволюции и миграциях до-сапиентных представителей рода Homo, которые, несмотря на очень скромные технические возможности и многочисленные природные препятствия, смогли занять ведущее положение в среде обитания, обойти всю Африку и пройти восемь тысяч километров по землям Евразии. Это было бесспорно большое достижение первых Homo, но огромные участки суши еще очень долго оставались безлюдными. В Америку и Австралию человек проник только на эволюционном уровне Homo sapiens.
Сейчас речь пойдет о части Света, открытие которой обычно считается заслугой Колумба или скандинавских викингов.
Первоначальное заселение Америки и происхождение коренного индейского населения — одна из «тем-долгожителей» в антропологии, археологии, истории. Ей ученые посвящали работы уже в семнадцатом веке. Например, в книге Н.Витсена, подготовку и перевод которой лично курировал Петр Великий, была изложена детально разработанная гипотеза переселения в Америку «азиатов» из Сибири, причем в доказательство были приведены данные из области этнографии и антропологии. В конце XVIII века президент США, первый американский археолог Т. Джефферсон также отстаивал идею азиатского происхождения американских индейцев. Вероятно, определенную роль здесь сыграл антропологический фактор — физиономическое сходство аборигенов Америки с сибирскими народами. Однако, не всех оно убеждало. В частности, создатель первой научной классификации французский ученый Ф. Бернье (1684 год) сближал америндов (так часто называют американских индейцев) не с сибирскими монголоидами, а с европеоидами. В восемнадцатом веке К. Линней и И. Блюменбах, выделили коренное индейское население в отдельную расу. Так же поступили в девятнадцатом веке Ж. Ламарк, а в начале двадцатого века — итальянский антрополог В. Джуфрида-Руджери, затем — американские исследователи К. Кун, С. Гарн и Дж. Бердсел. В двадцатом веке преобладающей точкой зрения становится включение америндов в монголоидную расу, в чем сыграл роль авторитет американского антрополога А. Грдлички. Его концепцию в данном вопросе разделили и российские антропологи — А. И. Ярхо, Г. Ф. Дебец, Н. Н. Чебоксаров. Возможно, эта доктрина в расоведении и определила взгляд антропологов на предполагаемые пути первоначального заселения Америки: в науке закрепилась т. н. «берингоморская» теория, согласно которой первые переселенцы пришли в Новый Свет через область Берингова пролива. Эта идея хорошо соответствовала не только данным расоведения, но и «географической логике».
Представление о единой волне заселения Америки через Берингов пролив определило еще одно широко распространенное вплоть до наших дней мнение, что аборигены Америки представляют единую в антропологическом отношении группу и имеют общий физический тип. Этот вывод, впрочем, был не нов и восходил к восемнадцатому веку, когда испанский путешественник и исследователь Антонио де Ульоа произнес слова, ставшие потом поговоркой: «Если увидишь одного индейца из любого региона, можешь сказать, что видел их всех» (Comas, 1966, р. 567). Возможно, это убеждение базируется на известном психологическом феномене, когда поверхностный взгляд наблюдателя, находящегося среди представителей иного этноса, приводит к заключению, что «все они на одно лицо». В этом направлении, конечно, на серьезной научной основе, развивалась деятельность последователей школы А. Грдлички, который сформулировал понятие «американский гомотип» (American Homotype). Эту точку зрения поддержал известный английский антрополог А. Кизс, позиция которого была несколько смягчена признанием, что локальные вариации — от племени к племени и от региона к региону все же существуют, однако субстратом для них служит фундаментальное сходство, явившееся следствием происхождения от одной общей, небольшой предковой группы. На таких исходных положениях базировалось научное направление в антропологической американистике, которое получило известность как «монорасовое». Нужно заметить, что критерии «американского гомотипа» и «монорасовой» модели в подходе к изучению аборигенного населения Америки были все же довольно спорными: в их основе лежали в основном показатели пигментации и недоставало важных признаков черепа и скелета.
Многие исследователи-американисты высказывали сомнения относительно описанной выше модели «американского гомотипа» и происхождения коренного индейского населения Нового Света. По мере изучения последнего, особенно Южной Америки, все более выявлялась вариабельность физического типа америндов, которая стала основой для создания альтернативных гипотез, прежде всего учитывающих разнородный антропологический тип аборигенов. Возникли «полирасовые» модели, среди которых наибольшей известностью пользовалась гипотеза французского ученого Поля Риве, пришедшего к выводу, что индейское население сформировалось в результате нескольких миграций групп людей, принадлежавших к четырем различным расовым типам, проникшим в Америку или через Берингов пролив (монголоидный компонент) или через Тихий океан (австралоидный и малайско-полинезийский компоненты). П. Риве определенно указал на присутствие австралоидного (или, как часто говорили в то время, — меланезоидного) компонента у индейцев Южной Америки, что было подтверждено позже многими антропологами и доставило дополнительные аргументы в пользу существования т. н. «палеоамериканского типа» (тип Лагоа-Санта, «лагиды»).
Весьма оригинальную гипотезу первоначального заселения Нового Света предложил португальский этнограф А. Мендес Корреа, который считал, что первые люди могли достигнуть Америки через периферийную зону Антарктиды в то время, когда отступал ледяной покров на этом материке. Действительно, если посмотреть на карту, то путь от Тасмании до Огненной Земли представляется довольно коротким, во всяком случае короче, чем траектория через весь Великий Океан. Однако, доказательств возможности перехода Антарктиды в эпоху верхнего палеолита никто привести не может и, скорее всего, не сможет: ледяной щит покрыл Антарктиду еще в миоцене, а в предполагаемый период описываемой миграции вряд ли были существенные потепления, судя по тому, что в северном полушарии вюрмские зимы были очень суровы. Правда, как сообщает Х.Комас, в Гренландии, например, между двенадцатым и пятнадцатым веками, по данным археологии, большой участок суши освобождался ото льда настолько, что там могли расти деревья. Кроме того, имеется и другая аргументация, в пользу гипотезы А. Мендес Корреа: краниологический тип «фуэгид» — коренных жителей Огненной Земли наиболее близок к австралийскому среди всех американских форм. При всем этом, однако, главная трудность для описанной гипотезы состоит в полном отсутствии археологических и палеоантропологических доказательств. В данной работе подобные предположения рассматриваются в основном с целью показать сложность проблем первоначального заселения Америки и многообразие подходов к их решению. Антропологические данные показывают необходимость особого подхода к изучению генезиса аборигенов Южной Америки, которые достаточно резко отличаются от своих северных соседей не только по физическому типу, но, возможно, и по происхождению и путям проникновения в Новый Свет. Большой неожиданностью может, в частности, оказаться для науки вполне вероятное заключение, что Южная Америка «открыта» человеком раньше Северной.
Чтобы наполнить приведенные выше рассуждения научными фактами, необходимо прежде всего обратиться к антропологической характеристике индейского населения Америки. В наши дни это сделать нелегко, т. к. европейская инвазия в Новый Свет очень сильно изменила исходную картину распределения племен и антропологических комплексов, сложившуюся в результате первоначального заселения и последующих внутриконтинентальных популяционных процессов.
Изменения, произошедшие в историческое время, наиболее сильно повлияли на физиономический тип населения. Поэтому исследования, выполненные в прошлом веке приобрели особую ценность, сохранив хотя бы часть первоначальной информации о внешнем облике племен, заселивших Новый Свет. Учитывая это, я в данной части биологической характеристики аборигенов Америки обращаюсь к трудам американистов прошлого столетия. В начале двадцатого века антропологические исследования в Америке проводили итальянские антропологи Дж. Серджи и Р. Биасутти. Несколько десятилетий спустя аналогичные работы выполнялись аргентинским ученым Х. Имбеллони и немецким исследователем Э. Эйкштедтом, которые систематизировали результаты работ своих предшественников и получили новые данные. В дальнейшем я буду в основном цитировать труды X. Имбеллони и Э. Эйкштедта, которые, что очень важно, получили сопоставимые и укладывающиеся в логичную схему описания морфотипов. Конечно, антропологические варианты, выделенные этими исследователями, в наши дни уже практически невозможно идентифицировать, но главное, что антропологи смогли показать гетерогенность коренного индейского населения в противовес идее «американского гомотипа».
Привожу вкратце антропологическую систематику доевропейского населения Америки по Х. Имбеллони (Imbelloni, 1938, 1943). В скобках даются названия выделенных вариантов по классификации Э. Эйкштедта.
1. Субарктиды (эскимосы). Аляска. Рост мужчин 158–164 — см., уменьшается с востока на запад. Короткие конечности, желтоватя кожа, довольно часто встречающийся эпикантус, мезо-долихокефалия (головной указатель 75–76) сильно выдающиеся скулы, средняя степень выступания носа, волосы черные, тугие, прямые.
2. Колумбиды (пацифиды). Группы северо-запада и севера тихоокеанского побережья. Рост высокий и средний (мужчины 161–170 см., женщины 152–160 см), выраженная брахикефалия (головной указатель 84–90), довольно светлая кожа, короткие ноги, слабое развитие третичного волосяного покрова.
3. Планиды (сильвиды). Занимали зону северной Америки от Аляски до Атлантического океана, ограниченную с севера тундровой зоной, а с юго-запада и запада — Скалистыми горами. Обитали в области бассейна реки Миссури. Высокий рост (мужчины 166–176 см., женщины 158 см), мезокефалия (головной указатель 79,3-81,5), выступающие скулы, сильно выступающий «орлиный» нос, волосы и глаза черные, цвет кожи светло-бронзовый. Именно этот тип, а также близкий к нему вариант «апаласиды», (см. пункт 11) фигурирует в художественной литературе как характерный облик «индейца».
4. Сонориды (маргиды). Занимают часть побережья Тихого океана, штаты Орегон и Калифорния, а также часть территории Мексики (штат Сонора). Рост довольно высокий (мужчины 166–170 см., женщины 152 см.), мезокефалия (головной указатель 78,5-80,0), малые размеры головы, узкий, наклонный лоб, сранительно темный (темнее, чем в группах 2–4) с красноватым оттенком цвет кожи.
5. Пуэбло-андиды (андиды). Расселены вдоль рек Рио Гранде, Колорадо, в штатах Аризона и Нью-Мексико. Известны как «индейцы пуэбло». На юге аналогичный тип локализуется в Андийской зоне Колумбии, Эквадора, Перу, Боливии, а также в северных районах Чили и Аргентины. Малый рост (мужчины 159–162 см.), мезо-брахикефалия (головной указатель 81,5-89,0), малые размеры головы, низкое лицо, нос широкий, при этом достаточно сильно выступающий, большой скуловой диаметр, варьирующий цвет кожи, преобладает интенсивная пигментация, волосы черные, жесткие, прямые, слабо развит третичный волосяной покров, часто встречается искусственная деформация головы.
6. Истмиды (централиды). Распространены от южной Мексики до Колумбии. Низкий рост (мужчины 150–158 см., женщины 143 см.), брахикефалия (головной указатель 86–89,5), лицо широкое и низкое, нос широкий (платириния), глаза черные, волосы черные, прямые, жесткие. Характерными представителями являются индейцы майя.
7. Амазониды (бразилиды). Занимают обширную зону Южной Америки от Анд до Атлантического океана, включая бассейны Амазонки и Ориноко, на юге — до реки Лa Плата. Рост средний (мужчины 155–158 см. на севере бассейна Амазонки, 161–166 см на юге). Сильно варьирующий головной укзатель (79–84), короткие ноги, длинные руки. Кожа различных оттенков на желтоватом фоне.
8. Пампиды (патагониды). Район Матто Гроссо, центральная равнина Аргентины, степи юга до Огненной Земли. Рост высокий и очень высокий: рост мужчин в области Чако 160–170 см., в Патагонии — 173–181 см. Брахикефалия у техуэльче (указатель 80,0), долихо-мезокефалия у она. Сильно выступающие скулы, крупные размеры лица, лепториния (относительно узкий и высокий нос), довольно темная пигментация кожи с бронзовым отливом, волосы прямые, жесткие, черные. По сообщениям аргентинских антропологов этот вариант антропологического типа в настоящее время уже не существует.
9. Лагиды (лагиды). Восточное плоскогорье Бразилии, несколько изо-лятов в других регионах, в частности — на крайнем юге Калифорнии. В прошлом этот тип был распространен гораздо шире — на территориях Мексики и Чили. Низкий рост (мужчины 150–157 см.), резко выраженная долихокефалия (головной указатель 66–73), высокий свод черепа, лицо широкое и низкое, относительно широкий и низкий нос, темная пигментация волос и глаз, форма волос и цвет кожи вариабельны. Этот вариант в значительной степени послужил антропополгам и, прежде всего, И. Деникеру, в начале двадцатого века основой для выделения т. н. «палеоамериканского» типа, происхождение которого вызвало долгую дискуссию о путях проникновения и физическом типе первых пришельцев на землю Нового Света.
9. Фуэгиды (лагиды). В антропологической классификации аборигенного населения Америки Э.Эйкштедт не выделяет аналогичного варианта и включает комплекс «фуэгид» в состав подразделения «лагиды». Основной ареал фуэгид — Огненная Земля, но отдельные очаги распространения этого типа отмечаются также на побережье Чили, среди некоторых племен Колумбии, в северной Калифорнии, среди ботокудов Атлантического побережья. В прошлом этот морфологический вариант был распространен среди многих ныне исчезнувших южноамериканских групп. Рост низкий (мужчины 157, женщины 147 см.), долихокефалия (головной указатель 73–77), узкий лоб, лепториния, сильно развитое надбровье. Вместе с лагидами этот вариант составлял основу «палеоамериканского» комплекса, характеризовавшего древнейшую волну заселения Америки.
10. Апаласиды (это понятие введено Х. Имбеллони позже) и обозначает индейское население зоны, прилегающей к атлантическому побережью Северной Америки. По большинству характеристик сходны с планидами (3), от которых отличается лишь выраженной долихокефалией. Характерные представители — гуроны (ирокезы) и алгонкины.
Классификация Х. Имбеллони (а, строго говоря, Серджи-Биасутти-Эйкштедта-Имбеллони) неоднократно становилась объектом критики. В частности, еще в 1951 году М. Т. Ньюмен (Newman, 1951) подверг сомнению концепцию Х. Имбеллони, считая ее «слишком миграционистской» в силу того, что в выделенных типах аргентинский автор видел отражение исходной политипии, многокомпонентности и признавал возможность сохранения исходных вариантов в разных экологических зонах в течение очень долгого времени. Сам М. Т. Ньюмен придерживался прямо противоположной, крайне «экологистской» позиции, полагая, что все разнообразие американских антропологических вариантов объясняется только влиянием локальных средовых условий. Позже эту же точку зрения разделил перуанский американист М. Валье (Valle, 1964). К сходным выводам относительно решающего значения гетерогенной среды в формировании антропологической политипии доколумбовой Америки, пришли также В. П. Алексеев (1974) и С. Молнар (Molnar, 1992). Но если американисты признали возможность столь быстрой адаптивной радиации исходных популяций, как же они так долго мирились с концепцией «американского гомотипа», согласно которой потомки одной небольшой группы мигрантов расселились по территории всего (!) Нового Света, не претерпев при этом сколько-нибудь существенных модификаций морфо-типа? Я склонен учитывать при анализе расогенетических процессов как средовые факторы, так и роль исходного генетического разнообразия, модифицированного и усиленного генным дрейфом в группах, оказавшихся в изоляции. Несомненно, критика приведенных выше схем антропологической классификации коренного населения Америки во многом справедлива. Нужно вспомнить, что эти работы выполнялись в тот период, когда в расоведении был принят типологический подход, который вызвал впоследстии бурю отрицательных эмоций и произвел решительный переворот в умах исследователей, быстро ставших на путь воинствующего популяционизма. Я не типологист и не апологет устаревшего типологического метода. Однако, при всех условиях считаю, что в любом случае нужно стараться найти «рациональное зерно» в добросовестных трудах наших предшественников. В данном случае этим рациональным зерном является констатация значительного разнообразия коренного населения доколумбовой Америки. Причины возникновения и сохранения этого разнообразия — другой вопрос, требующий, действительно серьезной дискуссии на современном уровне развития науки. Важно, что ученые двадцатого века, профессиональным взглядом осмотревшие громадные контингенты населения, не увидели в них никакого «американского гомотипа», но выделили некоторые неоднородности, представляющие интерес для антрополога. Классификация X. Имбеллони ярко иллюстрирует бросавшееся в глаза исследователя разнообразие не только в отношении черт большой монголоидной расы, но также по ряду признаков, выходящих за рамки этого комплекса, в частности не-монголоидных особенностей, составивших основу Аля выделения «палеоамериканского» комплекса. Они наблюдаются (или (наблюдались) чаще всего в периферийных регионах, концентрируясь преимущественно в группах, относимых X. Имбеллони к лагидам и фуэгидам, черты которых, поданным антропологов (см. выше) прежде были распространены гораздо шире, что само по себе наводит на мысль о древнем cjjoe и первичной волне инвазии человека в Новый Свет. В настоящее время антропологи все реже прибегают к измерениям и описаниям ныне живущего населения, так что информация по вышеизложенному разделу в общем существенно не обогащается. Зато наблюдается значительный приток новых данных по краниологии, одонтологии, дерматоглифике, генетическим маркерам. Переходим к описанию достижений краниологии в области проблем первоначального заселения Америки.
В этом вопросе наибольший интерес представляют краниологические материалы плейстоценового времени, т. е. древнее 10–12 тыс. лет. На территории Америки таких находок мало, притом их датировки постоянно пересматриваются и оспариваются. Хорошим примером этой ситуации является история с черепом из Отовало (Эквадор). Плейстоценовая датировка этой находки подверглась резкой критике (Brothwell & Burleigh, 1977) и, в конечном счете, даже сводилась по результатам коллагенового анализа к 2–3 тысячелетиям до нашего времени. Однако, проверка несколькими разными методами все же дала свидетельство значительной древности находки — до 28 тыс. лет (Davies, 1978), хотя эта дата до сих не является общепринятой. Согласно компромиссному мнению экземпляр в любом случае должен быть датирован концом плейстоцена (Rodriguez, 1987). В связи с предполагаемой значительной древностью черепа из Отовало заслуживают особого внимания его морфологические особенности: сильно развитый надглазничный валик, наклонный лоб, резкая долихокрания, крупные зубы. Автор находки Д. М. Дэйвис счел эти показатели достаточными, чтобы констатировать неандерталоидный облик черепа, однако по метрическим показателям экземпляр не выходит за пределы вариаций современных человеческих групп. Мне представляется, что комплекс Отовало скорее обнаруживает черты сравнительно недавно выделенного таксона Homo helmei — ранней суб-современной формы, которую авторы обычно не включают в Homo sapiens, считая пресапиентным звеном эволюции. «Типовым экземпляром» этого таксона называют череп из Флорисбада (McBrearty & Brooks, 2000). Рассматриваемый морфотип, с моей точки зрения, может быть также определен как «поздний архаичный сапиенс» по систематике Г. Бройера, что в общем мало меняет анатомический диагноз находки, но все же допускает включение последней в состав вида Homo sapiens. В докладе на международной конференции «Экология и демография человека в прошлом и настоящем» в 2004 году я высказал гипотезу, согласно которой «поздний архаичный сапиенс» должен рассматриваться как эволюционный пласт, явившийся основой поздней дифференциации человечества. Этот антропологический вариант широко расселился по всей Эйкумене, что доказывается находками позднеплейстоценовых черепов данной эволюционной стадии в Африке, Азии, Австралии и Америке. В частности, экземпляры Отовало, Пунин и Понтимело я считаю бесспорно принадлежащими к типу «позднего архаичного сапиенса», т. е., по более современной терминологии, — к Homo helmei (Зубов, 2004).
Д. М. Дэйвис дал описание еще одной интересной находки на территории Южной Америки — двух челюстей, обозначенных в каталогах как «Боливия-1» и «Боливия-2», датируемых концом плейстоцена и по морфологическим показателям также обнаруживающих черты архаизма. С моей точки зрения, они, вернее всего, также могут быть отнесены по эволюционному уровню к «позднему архаичному сапиенсу». Вообще плейстоценовые черепа с территории Нового Света неоднократно оценивались специалистами как «до-сапиентные» в силу их массивности, крайней долихокрании, сильной выраженности суперструктур, в особенности надбровья, альвеолярного прогнатизма, «килевидной» формы черепной крышки, крупных зубов. Однажды было даже сообщение о находке в Бразилии черепа Homo erectus (Bryan, 1978), однако череп был плохо датирован и вскоре утерян. Широко известная находка «Леди из Миннесоты» (череп девушки, датируемый верхним плейстоценом или ранним голоценом — 15–10 тыс. лет до современности) была оценена археологом Дж. Дженксом как «слишком примитивная, чтобы быть отнесенной к современному виду» (цит. по работе J. V. Rodriguez, 1987). Однако, большинство антропологов не согласились с этим и отметили лишь негроидный облик данного черепа, отличающегося резко выраженным альвеолярным прогнатизмом. Черты архаизма были зафиксированы на черепе из Мидленда (Техас), датируемом одними методами (радиокарбон) самым концом плейстоцена, а другими (уран) — более ранним временем (20 тыс. лет до современности), а также, — в той или иной мере, — на плейстоценовых калифорнийских экземплярах из местонахождений Тренквилити и Дель Мар (Ля Джолла), которые по результатам некоторых методов датировки относятся к очень древним эпохам (20–30 тыс. лет до современности). Для известного черепа из Юха (Калифорния) американский исследователь С. Роджерс определил возраст 21500 лет (Rogers, 1977). Ранние датировки калифорнийских палеоантропологических находок были подвергнуты сомнению и пересмотрены, как, например, было с материалами со стоянки Ля Джолла, однако за некоторыми все же сохранилась репутация очень древних. В частности, черепа из Лагуна Бич и Лос Анжелес по данным радиоуглеродного и коллагенового анализа определенно отнесли к плейстоценовому времени: абсолютная датировка — 17150 и 23600 лет соответственно (Plhak, 1980). По мнению антрополога Т. Д. Стьюарта (США) эти находки совершенно определенно принадлежат к современному виду Homo sapiens, причем не выходят за пределы вариаций американоидного типа. Т.Д.Стьюарт уточняет, что черепа из Лос Анжелес и Лагуна Бич сходны именно с локальным калифорнийским вариантом. Здесь важно помнить, что Калифорния входила в область распространения антропологических типов лагид и фуэгид, т. е. носителей палеоамериканского комплекса, т. е. здесь речь может идти о преемственности древних и современных форм. В этой связи нельзя не упомянуть также одну из работ известного исследователя Огненной Земли М. Гусинде (Gusinde, 1975), описавшего практически современный череп огнеземельца-алакалуфа, отметив при этом архаичные черты лагидоидного типа, в частности сильно развитый рельеф надбровья, большую толщину костей, гипердолихокранию, что наводит на мысль о связи с древними вариантами палеоамериканского типа.
Обращает на себя внимание несоответствие датировок археологических и антропологических материалов. Археологи обычно не признают дат, превышающих 13 тыс. лет, на чем основывается предполагаемое время первоначального заселения Америки человеком. К этому вопросу придется еще вернуться в данной главе.
Ископаемых палеоантропологических находок, относящихся к раннему голоцену, известно уже довольно много. Среди них скелеты с острова Санта Роса (Калифорния, 7–8 тыс. лет), Тепешпан (Мексика, 9-10 тыс. лет), Чимальуакан (Мексика, 10500 лет), Текендама, (Колумбия, 7–9 тыс. лет).
Комплекс «позднего архаичного сапиенса» очень ярко выражен в хорошо известной серии черепов из пещеры Сумидоуро, Лагоа Санта (Бразилия). В середине XIX века (1835–1844 гг.) в этой пещере вел раскопки датский археолог П. В. Лунд, которому удалось добыть богатый скелетный материал — останки по крайней мере 30 индивидуумов. Датировка находок по сообщению мексиканского антрополога Хуана Комаса (Comas, 1966) 10782±400 лет до современности. Новые недавние исследования (Neves et al., 2005) показали более позднюю и менее определенную дату — «>8000 лет»). Этой находке суждено было сыграть большую роль в исследовании истории заселения человеком Нового Света. Своеобразие краниологического типа найденных черепов побудило сначала некоторых антропологов выделить новую расу — «Лагоа Санта». Особенно необычными для Америки казались такие признаки, как гипсидолихокрания — удлиненная и высокая форма мозговой коробки, сильно развитые надбровные дуги, низкое лицо, низкие орбиты, прогнатизм. Этот комплекс навел антропологов на мысль о присутствии в древности в Южной Америке экваториального (австрало-меланезийского) компонента. Первым об этом заявил французский исследователь А. Катрфаж. Впоследствии французский этнограф и антрополог П. Риве (Rivet, 1957) построил рассмотренную выше поликомпонентную гипотезу происхождения индейцев Америки в результате четырех миграций, одна из которых, — водный путь через Тихий океан, — принесла, по мнению автора, в Америку австрало-меланезийский компонент. Опорой этой теории в значительной мере послужили черепа из Лагоа Санта, а также черты «палеоамериканского» антропологического типа, выделенного в конце XIX — начале XX в И.Деникером (Dertiker, 1902). Специально проведенное исследование показало, что ведущим маркирующим признаком этого типа на краниологическом материале является сочетание австрало-меланезийских черт с нешироким (менее 25 мм) грушевидным отверстием (Зубов, 1968, 1993, 1999), причем этот комплекс, названный мною лагидоидным по местнахождению Лагоа Санта и по наименованию соответствующего антропологического типа лагиды, имеет закономерную связь с периферийными и труднодоступными районами Америки (Калифорния, бассейн Амазонки, Огненная Земля), что говорит о его большой древности. Интересно далее, что такая периферийная (но поздняя) группа, как американские эскимосы, обнаруживает лагидоидные черты, вероятно вследствие того, что древнейшие группы предков эскимосов, проникшие в Америку, могли иметь контакт с палеоамериканским по антропологическому типу индейским населением (например, с древнекалифорнийскими группами, занимавшими более широкий ареал). Эти факты были положены в моих вышеупомянутых работах в основу гипотезы, согласно которой в эскимосских популяциях, занимающих периферийные регионы Нового Света, имеются элементы палеоамериканско-го комплекса, в какой-то мере связывающие поздние волны мигрантов с древнейшим местным субстратом. Наиболее серьезным возражением против этой постановки вопроса является ссылка на большую высоту лица на черепе эскимосов по сравнению с палеоамериканоидами, в частности — с огнеземельцами. В ответ можно напомнить, что в ископаемой серии эскимосов с рстрова Св. Лаврентия высота лица оказалась в среднем равной 73,5 мм, что в точности соответствует этому размеру у лагид и фуэгид. По остальным краниологическим параметрам американские эскимосы почти до идентичности сходны с фуэгидами-огнеземельцами. Учитывая эти данные, можно говорить о панамериканском масштабе распространения древнейших лагидоидных элементов, игравших субстратную роль по отношению к новым миграциям.
По имеющимся в настоящее время данным мы можем охарактеризовать палеоамериканркий тип, кое-где в Америке сохранившийся до наших дней и близкий к австрало-меланезийскому морфологическому варианту. Прежде всего в нем надо отметить сочетание гипсидолихокрании (узкой и высокой черепной коробки) с низким и нешироким лицом. Развитые надбровные дуги и альвеолярный прогнатизм еще более подчеркивают сходство с австронезийским комплексом. Однако, признавая отмеченные краниологические параллели, их все же нельзя абсолютизировать: в описаниях древних американских черепов мало внимания уделяется такому важному таксономическому признаку, как ширина носа, которую в палеоамериканских сериях, как было отмечено выше, можно признать скорее небольшой, точнее — от малой до средней, как правило не более 24 мм. В то же время, у современных австралийских аборигенов ширина грушевидного отверстия черепа в среднем равна 26,9 мм, а на ископаемых черепах из Кохуны и Кейлора она еще больше: 32,0 мм и 27,0 мм соответственно (Рогинский, Леван, 1963).
Колумбийский антрополог X. В. Родригес (Rodriguez, 1987), изучая черепные серии Америки докерамического и керамического периодов, пришел к выводу, что проамериканский комплекс не укладывается в рамки какой-либо одной из трех больших рас Homo sapiens, будучи в известной степени промежуточным между последними: от экваториальных форм он отличается узким, довольно сильно выступающим носом и меньшим прогнатизмом, от азиатских монголоидов, особенно сибирских, — меньшей уплощенностью лицевого отдела и небольшими размерами лица, что дает исследователю прр заключить, что морфотип америндов с древнейших
времен выделялся своеобразием, достаточным для возведения америка-ноидного расового типа в ранг крупнейших подразделений человечества. Это, кстати, соответствовало бы трем важнейшим критериям систематики — ареальному, морфологическому и показателю внутренней дифференциации таксона. Здесь, правда, нужно заметить, что различия между «большими» и «малыми» расами достаточно условны. Поэтому, рассуждая о внутривидовом разнообразии человечества, я предпочел бы употреблять термины «антропологический тип» и «группа антропологических типов» (Зубов, 1996).
X. В. Родригес, выводя тип «среднего палеоамериканца», ссылался не только на типичные лагидоидные формы, но и на все более или менее древние краниологические материалы, в результате чего итоговый усредненный тип сдвинулся в сторону промежуточных общеамериканских значений признаков: назомалярный угол — 136–142°, зигомаксиллярный — 126,5-128,6°, средние абсолютные размеры лица, умеренный прогнатизм, средняя ширина носа. «Чистый» лагидоидный комплекс показал бы большее сближение с австрало-меланезийскими вариантами и несколько иные средние показатели, в частности — по ширине носа. Но главное, что и этот отдельно взятый тип древних америндов не мог быть однозначно отнесен к одному из трех-четырех основных внутривидовых подразделений человечества.
Вопрос об антропологических компонентах, вошедших в состав американских аборигенов, является спорным уже в течение многих десятилетий, постепенно претрпевая изменения в принципах и подходах дискутирующих сторон. В 1930 году американский антрополог Э. А. Хутон (Hooton, 1930) на основании детального исследования серии черепов индейцев Пекос Пуэбло из штата Нью-Мексико выделил в составе этой группы три краниологических компонента: 1. псевдо-австралоидный (близкий к айнскому); 2. псевдо-негроидный; 3. тип «корзинщиков» (близкий к европеоидному средиземноморскому). На этом субстрате, по Э. А. Хутону, сложилась еще в Старом Свете палеоамериканская миграционная волна. Впоследствии в Америку проникла новая волна мигрантов — брахикефалов монголоидного типа. Позже Э. А. Хутон несколько видоизменил свою схему, сведя ее к двум компонентам — монголоидному и восточно-негроидному (т. е. австрало-меланезийскому). Нужно отдать должное американскому исследователю, которому, даже на том этапе развития антропологии, который сейчас считают «устаревшим», удалось увидеть основные черты, из которых сложился физический тип америндов и установить наличие компонентов, принесенных в Новый Свет двумя волнами заселения, одна из которых имела австрало-меланезийскую основу. Лишь совсем недавно краниологические исследования бразильских антропологов полностью подтвердили эти выводы.
В 1951 году американист-антрополог Дж. Б. Бердсел (Birdsell, 1951) выступил с гипотезой участия в формировании физического типа аборигенов Америки компонента «архаичных европеоидов», под которым автор подразумевал «амурский тип», близкий к айнам. Сходную точку зрения высказал и Г. Ф. Дебец: он предположил наличие у древних индейцев двух компонентов — монголоидного и австралоидного, близкого к айнскому (Дебец, 1959). Дж. Б. Бердсел в доказательство наличия «амурского» типа в составе аборигенов Америки привел антропологическую характеристику верхнепалеолитического черепа UC101 из Верхнего грота пещеры Чжоукоудянь (Китай). Своеобразный краниологический комплекс этого экземпляра, согласно мнению Дж. Б.Бердсела, трансформировался в «амурский» тип, черты которого передались палеоамериканскому варианту (Пунин в Эквадоре, южнокалифорнийские плейстоценовые черепа). Сравнение с черепом из Верхнего грота Чжоукоудянь представляется очень уместным и перспективным. Сложным оказался вопрос о датировке этой находки. Результаты разных исследований были противоречивыми: от 10 тыс. до 30 тыс. лет до современности. По самым последним данным древность черепов из Верхнего грота равна 10175±360 лет до современности (Neves et al., 2005).
В Верхнем гроте пещеры Чжоукоудянь были обнаружены три черепа: UC101, UC102 и UC 103, отчетливо различающихся по краниологическому комплексу. Первый особенно интересен в связи с проблемой заселения Америки: он отличается долихокранией (указатель 70,2), очень большой длиной и высотой мозговой коробки (204 и 136 мм соответственно), высоким (77 мм) и довольно широким (143 мм), ортогнатным (угол 84°) лицом, низкими орбитами (31,5 мм), широким носовым отверстием (32 мм). Лоб наклонный, с развитыми надглазничными дугами, лицо значительно менее уплощено, чем у современных монголоидов. Краниологический комплекс экземпляра UC 101 исключает возможность рассматривать его как монголоидный. Не случайно, что сопоставление этой находки с черепами трех основных современных подразделений человечества не выявило определенных параллелей с каким-либо из них, зато было обнаружено несомненное сходство с черепами полинезийцев и айнов, которых относят к числу промежуточных, «контактных» форм (Рогинский, Левин, 1978). В. В. Бунак и М. М. Герасимова констатировали значительную близость описываемого черепа к европейскому верхнепалеолитическому экземпляру Сунгирь 1 (Бунак, Герасимова, 1984). При этом исследователи пришли к выводу, что UC 101 имеет даже меньше монголоидных черт, чем его европейский «близнец» из Сунгирского погребения в центральной России. Этот череп многие антропологи считают примером «недифференцированного» или «генерализованного» типа, определить место которого в рамках имеющихся расовых классификаций практически невозможно.
Его относили к примитивным монголоидам, европеоидам, полинезийцам, американским индейцам, однако ни в одном случае не приводились достаточно убедительные доказательства.
Череп UC 102 крупный, длинный (196 мм), долихокранный (указатель 69,3), очень высокий (150 мм), с узким (131 мм), низким (69 мм), ме-зогнатным, сильно профилированным лицом (назомалярный угол 130°, зигомаксиллярный — 130°), умеренно широким грушевидным отверстием (26 мм). Выступание носовых костей слабое, лоб сильно наклонный.
Женский череп UC 103 меньшего размера (продольный диаметр 184 мм, поперечный 131 мм), долихокранный (указатель 71,3), высокий (143 мм). Лицо широкое (137 мм), низкое (68,5 мм), уплощенное в верхнем отделе (назомалярный угол 150°) и профилированное в средней части (127°). Дисгармония назомалярного и зигомаксиллярного углов на этом черепе выржена особенно резко. Носовое отверстие средней ширины (25,5 мм), орбиты низкие (31 мм). Сильно выражена «крышеобразная» форма свода мозговой коробки.
Ф. Вейденрейх сближал череп UC 102 с меланезийским типом, UC 103 — с эскимосским, полагая, что в этом регионе происходило интенсивное межгрупповое смешение. Здесь уместно вспомнить точку зрения В. В. Бунака относительно краниологического полиморфизма людей верхнего палеолита: сравнение трех описанных черепов представляет хорошую иллюстрацию этой гипотезы. Это был период недифференцированных, генерализованных краниологических комплексов, из среды которых выделились группы, заселившие Новый Свет.
В 2003 году американские антропологи Д. Л. Каннингэм и Р. Л. Янтц опубликовали результаты детального сравнительного исследования черепов UC 101 и UC 103 из Верхнего грота Чжоукоудянь. В результате статистического анализа выяснилось, что череп UC 101 обнаруживает большое сходство с полинезийской популяцией с острова Пасхи, а также, отчасти, — с европеоидами, в то время как UC 103 четко сближается с австрало-меланезийскими группами. Особенно важно при этом, что оба экземпляра имеют общие черты с палеоамериканскими краниологическими сериями. Изученные черепа не входят в состав каких-либо известных внутривидовых подразделений, но, вероятно, происходят из генерализованной популяции, которая была предковой для палеоамериканской волны заселения Нового Света (Cunningham & Jantz, 2003).
Еще один вриант классификации антропологических типов Америки был предложен В. П. Алексеевым, который выделил в составе аборигенного населения две основные расы: североамериканскую и центрально-южноамериканскую, в пределах которых констатирована дальнейшая дифференциация на группы популяций, формировавшиеся вокруг нескольких очагов расообразования (атлантическая, тихоокеанская, калифорнийская, центральноамериканская, амазонская, андская, патагонская, огнеземель-ская) (Алексеев, 1974). Этот автор предположил, что в ходе формирования расового состава аборигенного населения Америки определенную роль сыграл европеоидный компонент, проникший вместе с другими составляющими через область Берингова пролива. На это X. В. Родригес возразил, что европеоидный компонент в Азии вряд ли древнее 7–6 тыс. лет, т. е. он мог принять участие в формировании лишь сравнительно поздних комплексов. Можно также предположить, что европеоидные (или «псев-доевропеоидные») черты были связаны с общим недифференцированным элементом, входившим в состав протоамериканоидов. Гипотеза европеоидной примеси у индейцев Америки высказывалась и ранее: так, Хуан Комас (Comas, 1973) считал, что какие-то группы переселенцев из Европы могли проникнуть в Америку с востока, через Гренландию в периоды потепления климата или же через Атлантический океан, располагая уже некоторыми примитивными средствами навигации. Этот мексиканский антрополог допускал возможность миграции человека таким путем еще в мадленскую эпоху (10–15 тыс. лет назад), причем это событие могло, по мнению X. Комаса, даже в какой-то мере повлиять на физический тип населения. Мне представляется, что гипотезы, предполагающие участие европеоидного компонента в формировании антропологического типа индейцев в основном базировались на таких признаках (кстати, связанных значительной корреляцией) как выступание носовых костей и общая вертикальная профилировка лица. С моей точки зрения здесь мог иметь место случай гомоплазии (т. е. наличия сходного проявления признака в разных таксонах за счет механизмов, не связанных с общностью происхождения). В пользу такой точки зрения свидетельствует одонтологический материал: частота лопатообразной формы резцов у индейцев Северной Америки соответствует мировому максимуму, в то время как у европеоидов отмечены как раз минимальные показатели концентрации этого очень важного дифференцирующего внутривидового признака.
В 1937 году Я. Я. Рогинский выдвинул и обосновал гипотезу заселения Америки популяциями-носителями несформировавшегося (по параметрам современной внутривидовой таксономии) комплекса, характерными чертами которого по сравнению с современными монголоидами были: меньшая уплощенность лица, сильнее выступающий нос, высокое переносье. Важно отметить, что Я. Я. Рогинский приводил в качестве примера такого «недифференцированного» морфотипа упомянутый выше череп UC 101. Исходя из особенностей морфологии последнего, можно было бы добавить такие признаки, как выраженная гипсидолихокрания и сильно развитые надбровные дуги, т. е. те черты, которые сразу же напоминают нам лагидоидный палеоамериканский комплекс. По Я. Я. Рогинскому современный монголоидный тип с уплощенным лицом и эпикантусом сложился сравнительно поздно, причем в Восточной и Юго-Восточной Азии ему предшествовали «нейтральные», генерализованные варианты. Практически такой же точки зрения придерживался американский антрополог У.Хауэлз (Howells, 1940), полагавший, что первоначальное заселение Америки имело место в эпоху господства в Восточной Азии «мало специализированных» типов, дальнейшая эволюция которых привела к становлению современного монголоидного комплекса. Слабая выраженность последнего на древних черепах, помимо рассмотренного выше UC 101, явилась причинаой того, что исследователи, занимавшиеся палеоантропологией Америки, могли констатировать на данном материале лишь принадлежность его к «протоиндейским монголоидам» («Леди из Миннесоты» по Дж. Дженксу), либо к «протомонголоидам» («Трэнквилити» по Дж. Энджелу), а чаще просто ссылались на слабую выраженность монголоидных черт, как, например, поступил С. Роджерс при описании древнекалифорнийского черепа из Юха. Неспециализированный в отношении внутривидовой дифференциации характер проявляется в отдельных случаях даже в некоторых группах современного коренного населения Восточной и Юго-Восточной Азии, по ряду признаков сближающихся с америндами (седанги и баанары по Я. Я. Рогинскому).
Чем древнее азиатская форма, тем менее четко выражен у нее монголоидный краниологический комплекс и отчетливее просматриваются черты сходства с австрало-меланезийскими вариантами. Индонезийский антрополог Т. Джекоб (Jacob, 1967) проследил последовательность микроэволюционных изменений древнего населения острова Бали, причем обнаружилась определенная тенденция к «монголизации» в направлении к современности и «австрализации» по мере увеличения древности человеческих останков. Более широкое расселение в древности австралоидов, точнее австрало-веддоидов, доказывается также сохранением черт данного типа в ряде современных групп Камбоджи, Вьетнама, Индии. В моих прежних работах (Зубов, 1968, 1979, 1993) не раз высказывалась мысль, что Новый Свет заселялся последовательными волнами мигрантов, каждая из которых отражала определенную стадию формирования монголоидного типа и дифференциации первичного общего «восточного» австрало-монголоидного ствола. При этом, как это было в Юго-Восточной Азии, высокий удельный вес австралоидного комплекса был характерен для относительно более древних периодов эволюции последнего. Именно этот протоморфный вариант был характерен для «первооткрывателей» Америки — носителей палеоамериканского физического типа. Продолжающийся поступательный процесс дивергенции монголоидной ветви «восточного надрасового ствола» обусловил возрастающую «монголизацию» последующих групп переселенцев, проникавших в Америку, так что более поздние волны приобретали более выраженный монголоидный облик, а более ранние сохраняли в себе много протоморфных черт австрало-меланезийского характера.
Прогрессирующую «монголизацию» мигрантов в Америку отметил в своем обширном краниологическом исследовании X. В. Родригес (Rodriguez, 1987). Согласно вычислениям этого ученого удельный вес монголоидного компонента в краниологическом (фенотипическом) комплексе палеоамериндов во время первоначального заселения Америки равнялся примерно 62,6 %, что аналогично показателям «монголоидности» уральского антропологического типа. Более поздняя волна мигрантов, — предков племен языковой семьи на-дене, — имела уже более высокий «уровень принадлежности» к монголоидному комплексу (85,7 %); наконец у самых поздних мигрантов (эскимосов, алеутов) доля этого компонента приближается к 100 %. Конечно, эти цифры в сильной степени зависят от набора признаков и выбора групп, но все же они в какой-то мере неплохо иллюстрируют поступательный характер специализации монголоидной ветви «восточного» надрасового ствола. X. В. Родригес приходит к выводу, что вариабельность америндов, особенно центральных и южных, имеет в своей основе полиморфизм палеоамериканского субстрата, где монголоидные и австралоидные элементы были слиты в едином корне. В ходе дифференциации в гетерогенных природных условиях Америки, благодаря селективным процессам, и генному дрейфу возникали локальные тенденции к усилению тех или иных особенностей комплекса, создавая впечатление значительного разнообразия.
X. В. Родригес детально изучил явления эпохальной изменчивости, — секулярного тренда, — в Америке, причем убедительно показал, что в ряде районов Нового Света, начиная с III–I тысячелетия до н. э., и в особенности после X–XI вв н. э., происходил быстрый процесс брахикефализации, так что без какого-либо притока инородных генов черепной указатель быстро менялся: долихокранные формы сменялись мезокранными и далее — брахикранными. Автор объясняет этот феномен переходом к оседлости и новому типу хозяйства — земледелию, с чем связана «пищевая революция» и соответственно — ослабление нагрузок на зубо-челюстной аппарат и перестройку отделов черепа, связанных с жевательной мускулатурой.
С учетом рассмотренных выше факторов внутривидовой дифференциации X. В. Родригес выделяет в составе аборигенного населения Америки к моменту прихода европейцев два основных морфологических комплекса, аналогичные тем, которые описаны в упомянутой работе В. П. Алексеева (1974): североамериканский и центро-южноамериканский, разграничительный рубеж между которыми проходит примерно между тридцатым и тридцать пятым градусами с.ш. Североамериканский комплекс подразделяется на несколько вариантов: 1. циркумарктический (эскимосы, алеуты); 2. тихоокеанский, включающий языковую группу на-дене (атапаски, тлинкиты, хайда, апачи, пуэбло); 3. североатлантический (алгонкинский) (сиу, черноногие, понка, арикара, гуроны, чейены). Центро-южноамериканский комплекс также делится на варианты: 1. центральноамериканский (часть индейцев южных районов США); 2. циркумкарибский (индейцы с территорий северной Колумбии, Венесуэлы, Гайаны, Суринама, Антильских островов); 3. аборигены андийских регионов Колумбии, Эквадора, Боливии, Перу, Чили; 4. аргентинская пампа, Огненная Земля, часть Чили, Боливии, бассейна Амазонки; 5. амазонский вариант, материал по которому очень скуден, так что можно судить лишь о гетерогенности популяции и притоке генов из циркумкарибского, андийского и южного регионов (Rodriguez, 1983; цитируется по работе того же автора 1987 г.). Дополнительный анализ показал, что центро-южноамериканский комплекс обнаруживает хорошо выраженные черты палеоамериканского типа. Однако, в среднем тип америндов наиболее близок к монголоидам, хотя можно констатировать и некоторый уклон в сторону других антропологических вариантов, прежде всего — полинезийцев. Здесь нужно заметить, что в описываемой работе близость аборигенов Америки к монголоидам несколько преувеличена, т. к. в изученном X. В. Родригесом материале преобладали серии, относящиеся к сравнительно поздним волнам миграций, уже значительно «монголизированным». Если бы для сравнительного анализа были взяты только палеоамериканские экземпляры, результат мог быть иным.
В конце прошлого века начали раскопки бразильские антропологи, среди которых выделилась группа которую обычно возглавлял У. А. Невеш. Фактически работы этих исследователей ознаменовали новый этап изучения процессов формирования аборигенного населения Америки. В 1989 году были опубликованы результаты обработки нескольких древних черепных серий (Текендама из Колумбии, черепа из нескольких могильников Бразилии, относящиеся к концу плейстоцена — началу голоцена) (Neves, Pucciarelli, 1989). Анализ древнейших черепных серий четко показал наибольшую близость палеоамериканских выборок к австралийским аборигенам, тасманийцам, меланезийцам. Авторы пришли к выводу, что предками палеоамериканских групп Америки были племена, относящиеся к генерализованному экваториальному типу, который был распространен в Азии до формирования монголоидов. Краниология т. о. не дает основания для включения всех америндов в состав монголоидной большой расы. Можно лишь говорить о родстве с последней лишь на уровне древнего общего протоморфного ствола. Австралоидные черты наиболее отчетливо проступают в морфологии древнейших палеоамериканских черепов.
В 1993 г. в очередной публикации группа У. Невеша подтвердила свои прежние выводы относительно домонголоидной волны заселения Америки (Neves et al., 1993).
К такому же выводу пришла американская исследовательница-палеоантрополог M. Лap,которая в своей работе 1995 года (Lahr, 1995) определенно показала, что первые обитатели Нового Света были не монголоидами, а австралоидами. В моих работах (Зубов, 1977, 1993, 1999) я проводил мысль о том, что обе эти расы являются результатом дифференциации первичного общего австрало-монголоидного ствола — первоначального внутривидового подразделения Homo sapiens. При этом я добавил бы, что итоговый вариант антропологического типа америндов включил, кроме того, некоторые изменения, возникшие в результате долгой изоляции в условиях Америки, так что я согласен с Хуаном Комасом (Comas, 1966, op. cit.), который писал, что в антропологическом отношении тип америнда есть нечто новое.
В 1998 г. группа У Невеша опубликовала очень важные данные о найденных в местонахождении Лапа Вермелья (Lapa Vermelha IV) в районе Лагоа Санта скелетах (42 мужских, 39 женских), датированных разными периодами времени, в основном самым началом голоцена. Среди них был череп молодой женщины в возрасте около 20 лет, получившей у археологов имя «Лусия» и приобретший широкую известность как своего рода символ прошлого Южной Америки. Знаковый характер этой находки объясняется ее большой древностью - 12,5 тыс. лет. Авторы публикации считают этот экземпляр не только древнейшим в Южной Америке, но и самым древним во всем Новом Свете (Neves et al., 1998). Как известно, далеко не все антропологи могут с этим согласиться (Irving, 1976, Зубов, 2004). Сравнительный краниологический анализ позволил сделать вывод, что найденный череп обнаруживает наибольшую близость к экземпляру № 103 из Верхнего Г пота пещеры Чжоукоудянь, а также к североафриканскому черепу Тафоральт 18. Исследователи приходят к общему выводу, что популяции, осуществившие первоначальное заселение Америки, были близки по типу к недифференцированному населению Азии, обитавшему в этой части Света до прихода монголоидов.
Бразильские исследователи, продолжают раскопки, дающие новые интересные археологические и антропологические материалы. В 2003 г. была опубликована работа, посвященная изучению скелетных останков людей из Сантано ду Риашо (центральная Бразилия) — примерно 40 индивидуумов (Neves et ai., 2003). Это место привлекло внимание археологов, т. к. там были обнаружены древние наскальные рисунки. Первоначальная датировка, произведенная методом радиоуглеродного анализа, составила 11960±250 лет до современности. В данном месте раскопок было отмечено наличие еще долее раннего слоя, содержащего древесный уголь, что могло указывать на присутствие человека. Однако, в этом вопросе исследователи проявили особую осторожность, полагая, что древесный уголь, по которому Цределялась древность местонахождения, мог иметь
естественное происхождение. В конечном счете авторы пришли к заключению, что изучаемый регион был заселен человеком («палеоиндейцами») в период между 9500 и 8200 лет до современности. Найдены орудия из кварца и следы охры. Из всего костного материала оказалось возможным выделить только шесть черепов, пригодных для измерений. Черепа характеризуются большой длиной и высотой, малой шириной мозговой коробки, низкими и узкими лицами, низкими орбитами. Многомерный анализ показал, что краниологический тип находок определенно сближает последние по всем основным показателям с современным коренным населением Австралии и Африки, не обнаруживая никакого сходства с североазиатскими группами и даже с современными индейскими популяциями Северной Америки. И в данном случае приходится вспомнить факты близости палеоамериканских краниологических типов с экземплярами из Верхнего грота пещеры Чжоукоудянь. Так, сопоставление черепа SR III (Сантана ду Риашо) с известным черепом UC 101 из упомянутой пещеры дает картину явного сходства обеих находок. Это становится особенно наглядным при наложении контуров мозгового черепа, которые практически совпадают у сравниваемых экземпляров (рис. 9).

Сопоставление черепов Сантано ду Риашо (1) с экземпляром и С101 из Верхнего грота Чжоукоудянь (2). Обвод мозгового черепа UC 101 вписан в обвод SR III (Сантано ду Риашо). Масштаб не соблюдается. Меняя размеры можно добиться практически полного совпадения контуров.
В 2004 году бразильские антропологи опубликовали новое исследование по краниологии древнего населения Южной Америки. (Neves et al., 2004). На этот раз речь шла о черепе из раннеголоценового местонахождения Капелинья II (Capelinha II), локализованного недалеко от Атлантического побережья, в долине Рибейра ду Игуапе (юго-восточный регион штата Сан Пауло, южная Бразилия). В районе Капелинья обнаружено 6 палеоиндейских погребений, но краниологический материал получен только из одного. Датировка находки — 8860±60 лет до современности. Череп Капелинья II мужской, средней степени сохранности. Он характеризуется довольно большой длиной (180 мм), сравнительно малой шириной мозговой коробки, узким (130 мм) и низким (71 мм) лицом, средней высотой орбит (35 мм), небольшой шириной грушевидного отверстия. Произведен полный комплекс обычных («классических») измерений и геометрический морфометрический анализ. Результаты сопоставлены с древнеиндейскими группами Южной и Центральной Америки, а также с различными по краниологическому типу сериями, опубликованными в работах У. Хауэллса. Рассматриваемый череп близко сходен с палеоамериканскими экземплярами, а также с черепами австралийцев, меланезийцев и современных негроидов Африки, что, по мнению авторов, подтверждает палеоамериканский статус находки. Нужно отметить, что в настоящее время термин «paleoamerican morphological complex» прочно вошел в лексикон антропологов, хотя несколько десятилетий назад само существование такого морфологического варианта ставилось под сомнение.
В 2007 году У. Невеш вместе с двумя другими бразильскими коллегами (Neves, Hubbe, Piló, 2007) опубликовали статью, в которой вновь обратились к материлам из Сумидоуро чтобы исследовать их на новом, более современном уровне, Интерес к этой черепной серии, открытой еще в середине XIX, понятен: Сумидоуро-Лагоа Санта — самая большая в мире коллекция палеоамериканских черепов, происходящих из одного места раскопок. Авторы применили шесть различных статистических методов анализа, причем привлекли обширный сравнительный материал из самых различных регионов Земли, включая, конечно, в первую очередь Америку. Результаты отчетливо показали сходство Сумидоуро с австрало-меланезийскими выборками при резком отличии от поздних (монголоидных) серий Азии и Америки. Полученные данные свидетельствуют в пользу гипотезы заселения Цового Света по крайней мере двумя сильно различающимися в биологическом отношении волнами мигрантов.
По мнению бразильских исследователей появление в Америке древнейших палеоамериканских популяций вполне можно объяснить не привлекая гипотезу трансокеанских миграций: в позднем плейстоцене в Азии был широко распространен краниологический тип, близкий к палеоамериканскому, и именно он был Принесен в Америку через Северо-Восточную Азию.
В недавно опубликованной работе бразильские антропологи анализируют предполагаемый процесс заселения Америки после «исхода» из Азии через область Берингова пролива (Pucciarelli et al., 2008). Выдвигается гипотеза, согласно которой тихоокеанский берег Северной Америки послужил «узким, но экологически непрерывным коридором», который прежде всего был освоен палеоамериканскими группами (op. cit., р. 305). Авторы ссылаются на работу американского археолога Э. Диксона, который предположил, что мигранты следовали не только «географической логике», но точнее — «географо-экологической логике» при выборе направления движения. Не могу не напомнить, что сходные принципы, включая гипотезу береговой адаптации и распространения вдоль морского берега, я положил в основу построения гипотетической картины «Великой миграции» с запада на восток Евразии. Э. Диксон предполагает, что береговой путь первоначальной дисперсии палеоамериканских групп обеспечивал достаточно богатый источник питания (включая «дары моря»), а также высокую скорость движения групп людей. Отсюда — генный поток в прибрежной зоне был более интенсивным, так что палеоамериканские черты могли быстро и далеко распространяться по краю континента. Это объясняет краниологическое сходство некоторых современных тихоокеанских групп, таких как Перику (Репей) с палеоамериканскими сериями. Тихоокеанский путь расселения («Pacific dispersal rout») помимо скорого передвижения по линии север-юг давал также возможность заселения прилежащих территорий: каждый его участок при определенных условиях мог стать стартовой площадкой для освоения внутренних районов континента. Авторы рассматриваемой работы анализируют дифференциацию индейских популяций по направлению запад-восток, причем различия между западными и восточными антропологическими вариантами отмечаются как в Южной, так и в Северной Америке. Береговая миграция от северной до южной оконечности Нового Света должна была занять примерно 12 тыс. лет. В пределах берегового коридора распространения древнеамериканских мигрантов краниологическая вариабельность была менее значительной, чем к востоку от Кордильер, где модель генного дрейфа по мере расселения играла все более важную роль в противоположность динамике изменчивости на западном побережье, которая сохраняла зависимость от фактора генного потока север-юг. Наличие двух различных ведущих факторов изменчивости приводила к дифференциации по направлению запад-восток в зависимости от соотношения сил генного потока и дрейфа (Pucciarelli et al., 2008). Такие же результаты, по свидетельству цитируемых авторов, были получены на основании генетического анализа по 16 системам. Известно также, что показатели социального и технологического прогресса к западу и к востоку от Кордильер различались по тому же направлению. По данным бразильских авторов в процессах дифференциации аборигенного населения Америки мог играть роль также селективный фактор, действовавший в ответ на гетерогенный характер новой для мигрантов среды. Однако, исследователи считают, что отбор не мог быть единственной силой, определяющей многообразие антропологических типов и придают очень большое значение генному дрейфу. Кроме того, бразильские антропологи, как и многие другие американисты, признают гипотезу исходной неоднородности антропологического состава мигрантов, предполагая, что заселение Нового Света происходило в два этапа, носивших гетерохронный и гетероморфный характер, причем первая, древнейшая волна принесла в Америку австрало-меланезийский антропологический комплекс, а со второй связана постепенная «монголизация» индейского населения.
В работе, посвященной исследованию скелетных остатков из Сантана ду Риашо (Neves et al., 2003) произведен детальный анализ вероятных путей дисперсии двух волн мигрантов на территории Америки и представлена карта, отражающая представление авторов о характере этого процесса. (Рис. 10). Предполагается, что отдаленные предки аборигенов
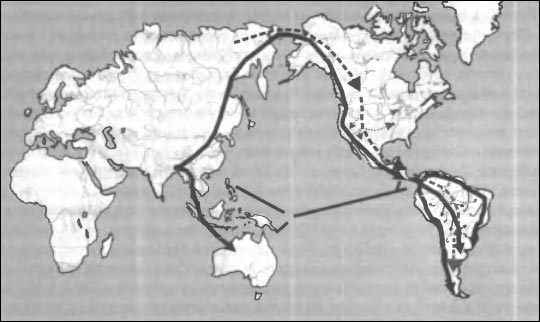
Предполагаемые пути заселения Америки. Гипотеза У. Невеша: траектория заселения через Берингов пролив показана черной линией (до-монголоидная палеоамериканская волна) и пунктиром (более поздняя монголоидная волна). Прямая линия через Тихий океан показывает путь миграции согласно гипотезе П. Риве и его более поздних последователей.
Америки вышли из Африки через Баб-эль-Мандебский пролив, следуя параллельно северному тропику дошли до территории Мьянмы, где пути мигрантов разделились: одна группа повернула на юг — в Индонезию, а затем в Австралию, другая ушла к северу, дошла до Берингии и южнее ледового щита переправилась в Америку. В Новом Свете основной путь мигрантов первой (австрало-меланезийской) волны лежал по «тихоокеанскому коридору» вплоть до Огненной Земли, но часть переселенцев обогнула Южноамериканский континент по северному и восточному берегам. Кроме того бразильские авторы считают возможным предположить, что палеоамериканская волна прошла также непосредственно с севера на юг в центральные области континента, распространяясь через тропическую сельву по притокам Амазонки и Ориноко. Это объясняет наличие «экваториальных» черт в краниологическом комплексе древнего населения района Лагоа Санта и центральной Бразилии. Продвижение древних популяций вдоль тихоокеанского и атлантического берегов позволяет понять причину сходства населения этих регионов в раннем голоцене. Авторы рассматриваемой статьи не согласны с довольно распространенной точкой зрения, согласно которой «экваториальные» краниологические особенности могли быть следствием естественного отбора в сходных климатических зонах Старого и Нового Света (не исключая полностью влияние этого фактора) и выражают уверенность, что австрало-меланезийский тип явился следствием сохранения «тропического комплекса», принесенного из Юго-Восточной Азии и, в конечном счете — из Африки. Сравнительно небольшие модификации, которые претерпел этот вариант древнего сапиенса (по Г. Бройеру — позднего архаичного сапиенса) в Новом Свете, связаны с быстрым продвижением палеоамериканских популяций по прибрежным зонам. Здесь уместно добавить, что скорость дисперсии вдоль берега могла быть увеличена за счет применения примитивных судов (Dixon, 1993). С моей точки зрения авторы вполне правы, когда проводят параллели между палеоамериканским типом и краниологическим вариантом Схул-Кафзех, полагая, что эти формы были звеньями в процессе эволюции и дифференциации древнего Homo sapiens, возникшего в Африке в глубокой древности — как теперь считают около 250 тыс. лет назад (по другим источникам — 130 тыс. лет назад). В это время появляется прогрессивная форма, которую одни исследователи относят к Homo sapiens, а другие выделяют в особый вид Homo helmei (McBrearty & Brooks, 2000), обладающий характерным набором краниологических черт, — долихокрания, сильное развитие надбровья, довольно низкое лицо, низкие орбиты, альвеолярный прогнатизм. «Типовым экземпляром» этого вида (по моему мнению — скорее подвида) исследователи считают африканский череп из Флорисбада, датировка которого (хотя и не бесспорная) — 250 тыс. лет до современности. К нему близки в морфологическом плане некоторые другие позднеплейстоценовые черепа Африки — Элие Спрингс, Летоли 18, Омо 2. Упомянутые авторы считают, что Homo helmei был непосредственным предком «настоящего» сапиенса, и отличался от последнего только некоторыми пережиточными чертами, постепенно устранявшимися микроэволюционными эпохальными изменениями, которые В. В. Бунак образно определил как «изживание архаичных особенностей, еще сильно заметных в верхнем палеолите» (Бунак, 1959, стр. 191). Если сделать мировой обзор краниологических материалов, относящихся к периоду поздний плейстоцен — ранний Полоцен, оказывается, что описанный выше комплекс черепа Флорисбад был широко распространен в ту эпоху и был одним из таксонов, вовлеченных в широкомасштабные мировые миграции. В частности, упоминавшиеся выше палеоамериканские черепа (Отовало, Пунин, Понтимело) характеризуются набором перечисленных выше черт Homo helmei. Те же особенности ярко выражены на ископаемых австралийских черепах (об этом — в следующей главе) и на китайском экземпляре из Дали, который, кстати, близок во времени к «типовому» образцу из Флорисбада. Создаётся впечатление, что в позднем плейстоцене началась глобальная дисперсия недавно возникшего нового подвида, в процессе которой был первоначально заселен Новый Свет и проникла очередная, сравнительно поздняя волна в Австралию. Этот динамичный таксон просуществовал более 200 тыс. лет и может считаться таксономически реальным звеном эволюционирующего рода Homo. Если мы вспомним теперь о «загадочном» палеамериканском типе, резонно будет искать его корни в числе потомков африканского Homo helmei. Конечно, все приведенные рассуждения строится на краниологическом материале, и мы с трудом представляем себе как выглядели живые «палеоамериканцы». Впрочем, некоторое предел Вление на этот счет могут дать современные австралийские аборигены, а также сохранившиеся кое-где в Америке группы «лагид» и «фуэгце» Огненной Земли, Калифорнии, бассейна Амазонки. Можно себе представить также, что такой же внешний облик имели представители «недифференцированных» домонголоидных популяций Юго-Восточной и восточной Азии, широко распространенных в конце плейстоцена. Дисг брсия Homo helmei и его прямых потомков может считаться третьей по счету межконтинентальной миграцией гоминин после первой «Великой» миграции и последующей волны расселения, связанной с распространением ашельской культуры, носители которой по антропологическому типу б.ч. относились к Homo heidelbergensis.
С моей точки зрения рассуждения бразильских коллег относительно формирования палеоамериканских антропологических комплексов в основном вполне логичны. Некоторые сомнения возникают в связи с трактовкой момента «исхода» третьей миграции из Африки 70 тыс. лет назад. Если «Флорисбадский» тип возник в Африке 250 тыс. лет назад и уже примерно 200 тыс. лет назад оказался в Китае (Дали), повторив в общем путь первой «Великой» миграции, то его носители должны были пересечь Баб-эль-Мандебский пролив не 70 тыс. лет назад, а значительно раньше. Флорисбадский комплекс, по мнению большинства исследователей, заслуживает названия «австралоидного», а не «афро-негроидного», от которого существенно отличается по строению лобной кости и надбровья. Поэтому, с моей точки зрения, нецелесообразно искать корни палеоамериканского типа среди представителей сформировавшихся негроидов Африки: более резонно прямо обратиться к древним африканским прото-австралоидам «флорисбадского» типа. Очевидно, инвазия палеоамериканоидов в Америку имела место через десятки тысяч лет после того, как сформировался и распространился в Восточной и Юго-Восточной Азии локальный пласт позднего «архаичного сапиенса», который может быть определен как «азиатский Homo helmei». По моему мнению, из этого пласта вышли популяции, осуществившие первоначальное заселение Америки и Австралии.
Продолжая описание фактического антропологического материала, на базе которого строится теоретическая модель процесса заселения Америки, перейдем к результатам исследований по программам одонтологии, дерматоглифики, генетических маркеров крови.
В работе американского антрополога А. Грдлички (Hrdlička, 1920) описывается т. н. «лопатообразная» (точнее — «совкообразная») форма верхних резцов, причем автор впервые использовал этот признак как таксономический критерий при сравнительном анализе сходства человеческих популяций. Как известно, лопатообразные резцы встречаются с очень большой частотой в монголоидных группах и у индейцев Америки (до 100%), в то время как в европейских и африканских популяциях они редки (до 0%), причем диапазон различий чрезвычайно велик и статистически вполне достоверен. Эти факты были рассмотрены исследователями как важный аргумент в пользу гипотезы принадлежности америндов к монголоидной расе и их происхождения из восточной Сибири. Таким образом первые одонтологические данные как будто подтвердили точку зрения А. Грдлички на антропологический состав аборигенного населения Америки и навели некоторых исследователей на мысль, что одонтологическая система в целом должна и далее «идти в ногу» с позицией этого исследователя, подтверждая, в частности, идею «американского гомотипа». В ходе дальнейших исследований в области одонтологии современного индейского населения Америки некоторые факты можно было рассматривать в русле гипотезы монголоидности индейцев. Так, в 1974 году я произвел выборочное обследование современного индейского и метисного населения тихоокеанского побережья Перу по полной принятой у нас одонтологической программе. Результаты этой работы были доложены на XIV Тихоокеанском конгрессе (Зубов, 1979). Сравнительный анализ продемонстрировал безусловную близость индейцев перуанского Побережья к дальневосточным монголоидам и монголам (халха и барга) по ряду признаков, кроме дистального гребня тригонида, частота которого оказалась у перуанцев намного ниже. Последующий анализ с использованием новых одонтологических критериев, включая одонтоглифику, показал, однако, что ставить знак равенства между перуанскими выборками и теми или иными североазиатскими популяциями по меньшей мере преждевременно. Так, по частоте мало изученного фена — переднего трансверсального гребня верхних моляров (часть гребня эпикриста) различия между индейцами и монголами (а также сибирскими монголоидами) очень велики: эпикриста встречается у перуанцев значительно чаще, чем в азиатских группах (Зубов, Халдеева, 1993). Заметим, кстати, что этот признак можно назвать скорее «экваториальным», чем «восточным» (монголоидным). Например, в Африке (население Мали) частота его в точности равна значению, отмеченному в Перу (35 %). Что касается сходства пёруанских индейцев с монголоидами по таким важным одонтологическим признакам, как лопатообразная форма резцов и коленчатая складка метаконида, то прежде чем делать на этом основании выводы о прямом родстве нужно учесть следующие обстоятельства: 1. монголоиды и австралоиды входят в единый «восточный» одонтологический комплекс, и провести дифференциальный анализ на присутствие того или другого компонента по отдельности затруднительно, тем более, что мы не располагаем подробными одонтологическими данными по австралийцам. Замечу здесь также, что результаты исследования австрало-веддоидных племен Индии, проведенного мною в 1968 году как раз свидетельствует о значительном одонтологическом сходстве последних с монголоидами, а это заставляет быть осторожным при трактовке «восточного» одонтологического комплекса, который может указывать как на монголоидность, — так и на австралоидность выборки; 2. наш материал по перуанским индейцам относится к Побережью, т. е. к зоне «Тихоокеанского коридора», в которую очень быстро проникали переселенцы «второй волны» — носители сравнительно высокой концентрации монголоидного компонента. Это предположение доказывается присутствием на севере Южной Америки такого антропологического типа, как «пуэбло-андиды» (по X. Имбеллони), проникшего из Северной Америки, вероятно значительно позже палеоамериканских мигрантов, по которым мы практически не имеем одонтологических данных.
Американский одонтолог К. Тернер (Turner, 1981, 1992) исследовал краниологические серии индейцев и эскимосов по своей палеоодонтологической программе, включающей наряду с общепринятыми критериями ряд признаков корневой системы, и пришел к выводу, что все аборигенное население Нового Света может быть разделено на три большие группы: 1. эскимосы-алеуты; 2. индейцы северо-запада Северной Америки; 3. остальные аборигенные популяции Северной и Южной Америки. Эти подразделения по К. Тернеру, отражают историю заселения Америки тремя волнами мигрантов: первая — более 15 тыс. лет назад, вторая — между 14 и 12 тысячелетиями до наших дней, третья — примерно 8500 лет назад. При этом не наблюдается четкого разграничения северного и южного одонтологических комплексов, т. е. не выделяется южноамериканский вариант, в котором, согласно многим исследованиям, должен наблюдаться максимальный удельный вес палеоамериканского компонента. Все изученные К. Тернером аборигенные группы Америки относятся к синодонтному типу, свойственному населению Северной Азии, Китая, Монголии, Северо-Востока Сибири. Альтернативный (сундадонтный) комплекс, по К. Тернеру, характеризует группы Юго-Восточной Азии.
Судя по периодически появляющимся публикациям различных авторов, одонтологическая карта Америки все же не выглядит столь монотонной. Так, по частотам важнейшего «восточного» (монголоидного) маркера — лопатообразной формы резцов, довольно гомогенным представляется только индейское население Северной Америки. В изученных различными исследователями группах индейцев пима, пуэбло, сиу, нол, в аборигенных группах Техаса, частота встречаемости этого признака колеблется от 90 до 100 %, однако в Центральной и Южной Америке дело обстоит иначе. Так, у индейцев кекчи из группы майя отмечено лишь 33,9 % лопатообразных резцов (Escobar et al., 1997), у мужчин племени гуахиро (Венесуэла) — 36,5 % (de Castillo, 1973). Эти цифры как раз соответствуют типу южных монголоидов, а также данным по полинезийцам, микронезийцам и айнам (ссылки см.: Зубов, Халдеева, 1989). Правда, в ряде южноамериканских групп процент рассматриваемого признака очень высок: у индейцев ленгуа (Парагвай) — 100 % (Kieser & Preston, 1981), у индейцев Певенче (Чили) — 95,3 % (Rothammer et al., 1968), у аборигенов северо-западной Аргентины — 84,0 % (Devoto et al., 1968). Можно было бы объяснить низкие частоты лопатообразной формы резцов в некоторых индейских группах европейской примесью (что, впрочем, противоречило бы сведениям, опубликованным упомянутыми исследователями), однако данные по достоверно метисным группам Америки не позволяют сделать такой вывод. Так, в смешанных выборках Северной Америки констатировано 85 % лопатообразной формы резцов, а в метисной группе Побережья Перу — 73 % (Зубов, 1979). Следовательно, речь идет о реальном разнообразии одонтологических комплексов Нового Света (Зубов, Халдеева, 1989). Это никак не согласуется с гипотезой «американского гомотипа» и скорее свидетельствует о неоднородном антропологическом составе миграционных волн, заселявших Америку в разные эпохи, причем указывает на наличие немонголоидного компонента в ряде групп мигрантов.
Некоторые исследователи предпочитали объяснять неоднородность одонтологического типа америндов эпохальной изменчивостью в пределах Нового Света. Однако, это может относиться только к абсолютным размерам зубов. Действительно, упомянутые выше проявления секулярного тренда в течение второго тысячелетия до н. э. в Америке отмечены также на одонтометрическом материале: даже в течение короткого периода — с 1600 до 1521 гг. до н. э. имела место редукция размеров зубов на 4,4 % (Christensen, 1998). Что касается деталей морфологии, их микроэволюция не зависела в такой степени от эпохальных факторов, связанных с переменами в образе жизни. Это доказывается тем фактом, что у азиатских народов, имеющих традиционно высокий уровень развития земледелия, например у китайцев, наблюдается концентрация лопатообразных резцов, близкая к мировому максимуму.
Свой оригинальный вклад в исследование разнообразия коренного населения Америки вносит также дерматоглифика. Имеются данные, согласно которым можно констатировать определенную однородность всего индейского населения: дельтовый индекс 13,4-13,7; индекс Камминса 7,2–7,5; узорность гипотенара 12–13 % (Хить, Долинова, 1990). Межрегиональная вариабельность наблюдается по узорности тенарной подушечки: здесь резко выделяются индейцы Центральной Америки. Североамериканские и южноамериканские группы практически не различаются. Даже такие своеобразные популяции, как огнеземельцы, в основном вписываются в общеамериканские параметры (Pereira da Silva, 1974).
По данным Г. Л. Хить и Н. А. Долиновой индейцы Америки в целом более близки к южным монголоидам, чем к северным (Хить, Долинова, 1990). Это особенно относится к группам Центральной и Южной Америки. В Северной Америке имеются популяции аборигенов, одинаково близких к южным и сибирским монголоидам. Эти факты противоречат концепции происхождения всех америндов от сибирских азиатских форм, а также позволяют считать коренное население Южной Америки близким к палеоамериканскому антропологическому комплексу. В упомянутой работе Г. Л. Хить и Н. А. Долиновой делается определенный вывод, что по сумме признаков дерматоглифики америнды как целое занимают промежуточное положение между монголоидами и австралоидами, причем те и другие в значительной степени сходны друг с другом. Предковый недифференцированный «восточный» комплекс просматривается здесь совершенно отчетливо.
Сходный вывод сделала группа американских антропологов, изучивших в 1979 г. ряд аборигенных групп Нового Света. Было констатировано промежуточное положение между монголоидами Восточной Азии и австралоидными вариантами (Garruto et al., 1979). Выявлена близость групп андийского региона, Амазонии и Центральной Америки, которые все вместе могут быть противопоставлены североамериканскому комплексу черт. Центральноамериканские популяции четко делятся на «майя» и «не-майя», а южноамериканские на «андид» и «не-андид». Эти данные показывают, что по вопросу о гомогенности или гетерогенности америндов по признакам дерматоглифики существуют разные точки зрения. Одни специалисты придерживаются взгляда на аборигенов Америки как на единый пласт населения, другие ставят под сомнение гипотезу единства типа америндов по данной системе признаков.
Выше мною была высказана мысль о том, что американские эскимосы не являются чисто инородным «клином» среди аборигенов Нового Света, но имеют в нем определенные древние корни, что можно показать на краниологическом материале. Данные дерматоглифики также дают основание для такого вывода: сравнительный анализ с учетом узорности тенарной подушечки позволяет поставить эскимосский комплекс между аборигенами Сибири и Северной Америки.
Было бы трудно делать итоговое заключение о формировании аборигенного населения Нового Света без данных по генетическим маркерам крови. Начавшиеся еще в первой половине двадцатого столетия исследования частот групп крови эритроцитарной системы АВО принесли чрезвычайно важные результаты, дающие представление о своеобразии генофонда америндов. Оказалось, что группа В практически полностью отсутствует у индейцев обеих Америк и крайне редка у эскимосов и алеутов, в отличие от сибирских монголоидов, где эта группа встречается достаточно часто. Следует, кстати, здесь же отметить, что аборигены Австралии, как и индейцы, лишены данного аллеля. В противоположность группе В другой аллель этой системы — О (нулевая или первая группа) обнаруживает диаметрально противоположную закономерность: у большей части североамериканских и практически у всех центрально-и южноамериканских индейцев частота этой группы достигает 90-100 %. Ее концентрация также высока у коренных жителей северо-востока Сибири, эскимосов, в ряде аборигенных групп Австралии и Океании. Интерес представляет и распределение группы А: у североамериканских индейцев и эскимосов она встречается относительно часто, но к югу — в Центральной и особенно Южной Америке ее частота снижается почти до нуля, так что в крови многих южноамериканских племен из системы АВО сохраняется лишь группа О.
Определенную информацию о генофонде американских индейцев дает система групп крови MNSs: частота группы М в Северной, Центральной и Южной Америке превышает 80 %, что является мировым максимумом, соответственно N очень редка по мировому масштабу. Аналогов такой структуры системы MNSs в мире нет. Достаточно яркую картину своеобразия аборигенного населения Америки дает система Rh (резус): случаи «резус-отрицательной» реакции у америндов крайне редки или вовсе не наблюдаются. Несомненный интерес в связи с темой данной главы представляет фактор Диего, найденный впервые в некоторых племенах Венесуэлы. Практически он может быть назван «южноамериканским маркером», т. к. только в этом регионе его частота достигает относительно высокого уровня (до 20 %), в то время как в Северной Америке этот признак встречается гораздо реже, иногда отмечается у азиатских монголоидов (особенно юго-восточных!), практически отсутствует у эскимосов, австралийцев, полинезийцев, а также в Европе и Африке (Garn, 1962).
В настоящее время очень большое значение придается изучению генетико-биохимической изменчивости человеческих популяций. Генетически детерминированные системы белков сыворотки дают в руки специалистов новые маркирующие показатели, применимые для анализа таксономических взаимоотношений различных групп человечества. Так, весьма интересна картина географического распределения гаплотипов иммуноглобулинов Gm, которую даже удалось связать с ареалами расселения трех больших рас (Спицын, 1985). В частности показано, что гаплотип, характерный для североазиатских монголоидов — нганасан и чукчей (Gm 1, 13, 15, 16) присутствует также у эскимосов и атапасков, т. е. представителей поздних волн заселения Нового Света и отсутствует в Южной Америке, где сохранилась варианты, общие для крупных подразделений человечества, т. е. маркеры недифференцированных комплексов (Сукерник, Кроуфорд, Осипова, Вибе, Шенфилд, 1988). Это, кстати, указывает на глубокую древность генотипов Южной Америки, представляющих палеоамериканскую волну.
На основе данных по полиморфизму групп крови и биохимических маркеров были сделаны важные выводы относительно антропологического состава америндов и их происхождения. Выявлены общие черты в генотипах аборигенов Америки и Сибири, причем особенно четко определилось сходство между индейцами и американскими эскимосами с одной стороны и коренным населением Северной Америки — с другой (Спицын, 1985; Сукерник, Кроуфорд, Вибе, Шенфилд, 1988; Quilici, 1977; Turner, 1992). Однако, это сходство в значительной мере относительно. Генетики М. Ней и А. Ройчоудхури, исследовавшие более ста генных локусов, констатируют генетическое своеобразие америндов и существенное различие между североамериканскими и южноамериканскими племенами. Выделяются эскимосы и атапаски Северной Америки как группы наиболее сходные между собой и наиболее близкие не к континентальным сибирским монголоидам, а к айнам (Nei, Roychoudhury, 1982). Генетическое разнообразие америндов оказалось более значительным, чем предполагалось. Особенно это касается населения Южной Америки. Так, венесуэльская исследовательница А. Г. де Диас Унгрия (de Diaz Ungria, 1966), изучавшая племена гуахиро, гуахибо, гуарао, яруро по системам эритроцитарных факторов крови (АВО, MNSs, Rh, Duffy, Diego), пришла к вывду, что перечисленные маркеры в сильной степени дифференцируют исследованные группы южноамериканских индейцев. Американский генетик Дж. Р. Нил, также посвятивший работу проблемам генетической гетерогенности южноамериканских племен, отметив выраженное своеобразие америндов в целом, подчеркнул, что дифференциация групп индейцев Южной Америки достигла очень высокой степени, для чего были необходимы чрезвычайно большие промежутки времени и жесткая изоляция. В частности, два племени, генетическое расстояние между которыми оказалось максимально большим, — яномама и шипибо, — должны были прожить в изоляции друг от друга по крайней мере 15 тысяч лет, чтобы возникли различия такого масштаба. На основании этих расчетов автор заключает, что первые мигранты должны были проникнуть в Южную Америку 20–25 тысяч лет назад, если, конечно, принять за основу гипотезу первоначального заселения Америки одной компактной волной мигрантов (Neel, 1973). Однако, как известно, эта гипотеза в настоящее время принимается все меньшим числом исследователей, причем существенную роль в этом играют генетики. Группа канадских генетиков во главе с М. Мансальве (Mansalve et al., 1994) пришла к выводу, что наиболее приемлемым объяснением разнообразия южноамериканских популяций может служить модель множества последовательных инвазий. Аналогичное заключение сделали бразильские исследователи (Bianchi et al., 1995). Таким образом, разнообразие аборигенного населения Америки, вероятно, является не только и даже не главным образом, следствием изоляции отдельных групп, расселившихся после первоначального проникновения в Америку какой-то одной популяции, но прежде всего — исходной генетической неоднородности групп мигрантов, пришедших в Новый Свет в разное время.
Здесь будет уместно подробнее остановиться на вопросе о времени первоначального заселения Нового Света, а также о путях проникновения первых людей в Америку, поскольку эти проблемы непосредственно связаны с основным содержанием данной работы.
Начну с рассмотрения обширного и разностороннего исследования, осуществленного группой американских генетиков из Университета штата Аризона (Williams et al., 1985). Эта группа ученых пришла к выводу, что в истории древнейших периодов заселения Америки можно выделить три периода — три волны мигрантов: палеоиндейцы (40–16 тыс. лет до современности); 2. группа индейцев, относящихся к языковой семье на-дене (14–12 тыс. лет назад); 3. эскимосско-алеутская волна (около 9 тыс. лет до современности). Аналогичная «трехволновая» модель принята также в работах К. Тернера, X. В. Родригеса, Д. Дьюмонда. Упомянутая российско-американская группа генетиков (Сукерник-Кроуфорд) занимает несколько иную позицию по рассматриваемой проблеме, считая, что было две волны первоначального заселения Америки: первая (палеоамериканская) волна — 20 тыс. лет назад, вторая (на-дене и эскимосы) 12-11 тыс. лет назад. Авторы не разделяют волны на-дене и эскимосско-алеутскую, полагая, что их дифференциация имела место уже в Новом Свете. Мне представляется заслуживающей внимания данная точка зрения, во всяком случае в том отношении, что она в какой-то мере включает американских эскимосов в круг «генетических америндов». Это, как говорилось выше, согласуется также с данными краниологии и дерматоглифики. Добавлю также, что отмеченный мною еще в 1968 году факт близкого краниологического сходства американских эскимосов с огнеземельцами наводит на мысль о возможности включения обеих этих групп в палеоамериканский антропологический вариант, представители которого по мере расселения новых мигрантов были оттеснены на периферию Америки. В свете описанных выше новых исследований бразильских антропологов огромное расстояние между ареалами расселения эскимосов и индейцев Огненной Земли не является основой для отрицания возможного родства двух рассматриваемых популяций, т. к. миграция в направлении Огненной Земли происходила быстро, следуя в пределах Тихоокеанского коридора. Скорость распространения групп людей в этом регионе увеличивалась, вероятно, также за счет использования навигационных средств, так что, по мнению Дж. Диксона, мигранты могли пройти по берегу путь от Аляски до Огненной Земли за 10 лет (Dixon, 1993).
«Двухволновая» гипотеза, как и «трехволновая» имеет своих сторонников (Quillici, 1977; Пласкетт и Диксон, 1979; Szathmary, 1985). Что касается времени первоначального проникновения людей в Америку, то широкий диапазон (40–16 тыс. лет до современности) по Р. Уильямсу или 34–17 тыс. лет до современности (Schurret al., 1993) привлекает во-первых осторожностью оценок и, во-вторых — признанием возможности очень ранней инвазии человека в Америку.
Высокий уровень генетической дифференциации популяций Южной Америки, также свидетельствует о необходимости основательно пересмотреть датировки и предполагаемые пути первоначального заселения Нового Света.
Древность первых людей в Америке, которая по определению многих американских археологов, не превышает 13–14 тыс. лет до современности, может оцениваться по-разному и оказаться значительно большей. Привожу ряд датировок плейстоценовых стоянок на территории Северной Америки (по Х. Комасу; Comas, 1966): пещера Пендехо (Нью-Мексико) — 39 тыс. лет; Луисвиль (Техас) — 38 тыс. лет; Сандиа (Нью-Мексико) — 30 тыс. лет; Америкен Фолс (Айдахо, 30 тыс. лет); Тьюль-Спрингс (Невада) — 28 тыс. лет, скальный навес Медоукрофт (Пенсильвания) — 19,6 тыс. лет.
В Южной Америке костатировано наличие столь же древних местонахождений: Монте Верде (Чили) — 33370 тыс. и Педра Фурада (Бразилия) — 32 тыс. лет (данные Dillehay & Collins,1988; Guèrin, 1993). Рассматривая приведенные цифры, хочется сразу же обратить внимание на тот факт, что они намного превышают принятые в настоящее время сроки первоначального заселения Америки и, кроме того, что они хорошо укладываются в период «взрывной» экспансии человека, начавшийся, по данным Дж. Диксона, в Восточной и Юго-Восточной Азии 40 тыс. назад. Полагаю, что он длился еще10-15 тыс. лет. Интересно добавить, что и в других частях Света, в частности в Европе, это также был период экспансии человека современного типа, вытеснявшего более древних гоминин.
В связи с оценкой древности эпизодов освоения человеком Нового Света возникает вопрос, действительно ли Южная Америка была заселена одной волной мигрантов в период «второй засухи» на Панамском перешейке, т. е. 15–13 тыс. лет назад. Эта гипотеза строится на том основании, что перешеек становился проходимым только когда пересыхали болота в тропических джунглях. Но ведь был еще период «первой засухи» около 30 тыс. лет назад, когда этим путем могли пройти более древние группы — носители палеоамериканского типа (Зубов, 1993).
Перечисленные выше древнейшие местонахождения следов пребывания человека могли бы резко переменить существующие в науке представления о ходе заселения Нового Света, однако этого не происходит отчасти в силу инерции общественного сознания, отчасти — в связи с оправданным желанием следовать лишь абсолютно достоверным фактам по принципу «подождем, посмотрим, а вдруг здесь что-то не так». Упоминавшийся мною известный американский археолог Дж. Диксон в своей книге «Quest for the origins of the first Americans» (Dixon, 1993) признает, что гипотеза проникновения первых американцев 14 тыс. лет назад хорошо объясняет дисперсию данной волны мигрантов, но не объясняет следы более раннего пребывания человека в Америке — 32 тыс. лет назад — Монте Верде (Чили), 33 тыс. лет назад — Педра Фурада (Бразилия), 17 тыс. лет (по другим источникам 19,6) — скальный навес Медоукрофт (Пенсильвания, Сев. Америка), датировки которых рассматривались как ошибочные (that have been regarded as equivocal — op. cit. p. 124). Меня всегда интересовало, в чем же здесь ошибка, и я не раз обращался с этим вопросом к археологам, но, к сожалению, не получил четкого ответа. Дж. Диксон в цитируемой работе от своего лица не называет эти датировки ошибочными, что видно, хотя бы из приведенной выше цитаты. Древнейшие даты принимаются им вполне серьезно, из чего следует признание, что Южная Америка могла быть колонизована раньше, чем Северная («исследователи должны быть готовы к такому выводу», стр. 132). Цитируя работу Д. Дж. Мельтцера, выдвинувшего концепцию «капельных миграций», Дж. Диксон соглашается с этим автором относительно того, что плейстоценовые группы, которым удалось пересечь Тихий океан, должны были быть очень малочисленными, и многие из них вымирали в силу невозможности освоиться в новой среде. Именно поэтому археологи находят так мало материальных свидетельств реальности тихоокеанского пути первоначального проникновения человека в Америку. Это в основном были ранние и неудачные попытки колонизации. В предыдущих главах мы рассматривали вероятность таких событий на других континентах. Можно думать, что они имели место гораздо чаще, чем нам теперь представляется. Дж. Диксон для примера напоминает о неудачно закончившейся инвазии викингов в Северную Америку. Подобные трагические эпизоды в истории заселения новых земель не вписывались в общую последовательность волн заселения Нового Света, так что ученые тщетно пытаются в каждом случае восстановить преемственность древнейших и более поздних капельных миграций. Закрепиться на новых территориях удавалось немногим. Но ведь не исключено и менее пессимистическое заключение. Палеоамериканская, австрало-меланезийская по типу группа смогла все же выжить и включить свои гены в смешанный генофонд коренного населения Америки, а кто был предком этих людей? Не те ли «герои тихоокеанской эпопеи», которые, по мнению некоторых современных исследователей, просто не существовали? В этой связи приходится вернуться к упоминавшейся в предыдущих главах важной проблеме — необходимости разграничивать два вида инвазии людей в неисследованные регионы: 1. первоначальное проникновение, которое могло заканчиваться возвратным движением, кратковременным, б. ч. сезонным, пребыванием (за которым следовало или не следовало закрепление), или, наконец, вымиранием; 2. колонизация — освоение территории, постоянное, длительное пребывание в течение всех времен года, сопровождающееся соответствующими модификациями моделей адаптации (как культурной, так и биологической). Я полагаю, что любая крупная инвазия человеческих популяций всегда начиналась с движения по первому типу, который являлся своего рода разведкой и прокладывал дорогу (иногда — ценою жизни многих людей) последующим, более многочисленным и «удачливым» мигрантам. Дж. Диксон говорит в своей работе в основном о колонизации земель, которая оставляла какие-либо следы, доступные для изучения методами археологии и антропологии. Но для той темы, которой посвящена моя работа, очень большое значение имеет первый, «разведывательный» этап инвазии, т. к. он характеризует готовность эволюционирующей группы человечества к следующему этапу прогрессивной деятельности, новому уровню возможностей человека в области познания мира и адаптации к неизвестным ситуациям. Этот период жизни человечества в разных регионах длился тысячелетиями. Он был труден для предков и труден для изучения в наше время, т. к. он оставил слишком мало абсолютно достоверной информации, чтобы войти в него как входит историк в летопись каких-либо средневековых сражений.
Важно, что Дж. Диксон не считает невероятным океанийский путь проникновения в Америку (правда, соответствующая глава его книги носит название «Speculations»). После плавания Т. Хейердала на плоту Кон-Тики трансокеанские путешествия с применением примитивных средств навигации стали казаться реальными. Оппозиционеры возражали, что норвежская экспедиция воспользовалась океанским течением, направленным из Америки в Полинезию, а не наоборот, т. е. путь в Америку был гораздо труднее. На это можно возразить, что противоположные течения в Тихом океане также существуют: взять, хотя бы, «Межпассатное противотечение», начинающееся близ Юго-Восточной Азии и доходящее до берегов Америки. Стоит теперь вспомнить, что период между 40 тыс. и 30 тыс. лет назад ознаменовался очень высокой, «взрывной» миграционной активностью, связанной несомненно с формированием «поведенчески современного человека» (behaviourly modern sapiens) и, судя по антропологическим и археологическим данным, положил начало новому этапу навигационных возможностей человека (см. гл. 3). Интересно сопоставить некоторые датировки находок по обе стороны Тихого океана: о-в Новая Ирландия (западная Океания, 31–33 тыс. лет назад) — Пещера Педра Фурада (Бразилия, 33 тыс. лет назад), пещера Монте Верде (Чили, 32 тыс. лет назад). Совпадение поразительное. Упомянутые стоянки, по мнению тех, кто не считает их датировки «ошибочными», являются древнейшими проявлениями оседлого образа жизни в Америке. Этот эпизод инвазии оказался тупиковым вследствие малого размера групп и слишком больших трудностей в ходе попыток освоения новой территории. Вероятно, эти «первые американцы» в конце концов погибли, но, может быть, оставили какой-то след в генофонде древнего, лагидоидного по типу южноамериканского населения. Дж. Диксон ставит в своей работе вопрос, могли ли люди достигнуть Америки только для того, чтобы погибнуть или быть поглощенными последующими волнами расселения и отвечает: «На основании моего опыта арктических исследований могу сказать — да!» и далее: «Материал по Австронезии показывает, что люди в Южном полушарии были способны предпринять морские путешествия 35–40 тыс. лет назад (Dixson, op. cit., p. 132).
Напомню также, что Мексиканский исследователь X. Комас не исключал возможности пересечения Тихого океана древними мореплавателями, причем приводил высказывания специалистов по направлению морских течении и ветров в пользу вероятности такого варианта океанической миграции (Comas, 1966).
Отрицая гипотезу пересечения океана с помощью очень примитивных навигационных средств, исследователи часто рассуждают с позиции среднего современного европейского человека, представляя себе какие чувства должен испытывать последний перед перспективой пройти Великий океан на маленьком суденышке. Здесь не учитывается психологический фактор: островитянин, выросший на островах Тихого океана, не боится океанских просторов, хорошо знает их и потому большей частью оказывается победителем в борьбе с этой понятной ему стихией. Тур Хейердал, проплывший на плоту от Америки до островов Туамоту, в книге «Аку-Аку» описывает эпизод из жизни полинезийцев острова Пасхи. Узнав об экспедиции Хейердала, эти люди решили повторить эксперимент: сколотили из досок маленькую лодку и пустились на ней в открытый океан. Пять недель спустя, они, голодные и измученные, но целые и невредимые, высадились на одном из атоллов Туамоту. У этих путешественников нашлись последователи: несколько жителей острова Пасхи на такой же маленькой лодке отправились в океан, имея только запас пресной воды и очень довольные высадились на берег Атиу, далеко к западу от Таити через несколько недель, пройдя морем тысячи километров (Хейердал, 1959). Надо сказать, что эти путешественники не имели ни карт, ни опыта далеких плаваний — их вели только интуиция и общее знакомство с морской стихией. Несомненно такими же были их верхнепалеолитические предки, которым был вполне по плечу вояж через весь Тихий океан до берегов Америки, тем более, что в плейстоцене этот путь был легче, чем теперь, благодаря снижению уровня мирового океана: было больше малых островов, многие из них были соединены сушей.
Упоминавшийся ранее французский ученый Ален Бомбар, прошедший Атлантический океан на маленькой резиновой лодке, рассказывал в своей книге, что после своего путешествия перестал бояться океана и считать его враждебной для человека стихией. В океане гибнет тот, кто боится.
Конечно, приведенные выше рассуждения не направлены против хорошо аргументированных гипотез заселения Америки через Берингию. Я просто хочу показать, что формирование антропологического типа американских аборигенов было сложнее, чем представлялось согласно гипотезе «американского гомотипа» и, возможно, включало элементы океанийского происхождения, проникавшие не только берингоморским путем, но и непосредственно со стороны Тихого океана.
Бразильские исследователи во главе с У. Невешем предпочли постулировать модель проникновения австрало-океанийских элементов через Берингию в составе древнейшей волны заселения (они придерживаются двухволновой концепции). В моих предыдущих работах (Зубов, 1999, 2002) я также допускаю возможность проникновения экваториальных элементов через Берингию в те времена, когда переселенцы характеризовались в основном австралоидным антропологическим комплексом, а монголоидный еще не сформировался. Отношение к океанийским миграциям у ученых могло, конечно, сложиться под влиянием критики, которой подверглась гипотеза П. Риве. Будущее покажет, кто прав, но в наше время необходимо помнить о таких находках, как Монте Верде и Педра Фурада и не торопиться без достаточно веских оснований списывать их со счетов, как «ошибочные».
Когда речь идет о числе волн мигрантов, проникавших в Америку, обычно подразумевается модель «одна волна — одна популяция». С моей точки зрения мы имеем право судить лишь о числе промежутков времени, в течение которых открывалась возможность «исхода» с территории Старого Света: о числе групп и тем более об их составе и численности мы ничего не знаем. Ведь эти периоды исчислялись не днями, а столетиями и даже тысячелетиями. Неужели за это время через Берингию успевала «проскочить» лишь одна гомогенная группа? Мне представляется более реалистичным допустить возможность проникновения в каждый период «зеленого света» нескольких групп, имевших разный антропологический тип. Тогда нужно говорить не о двух или трех гомогенных волнах, а о двух или трех миграционных потоках, включавших несколько гетерох-ронных и гетероморфных групп. Кажда волна должна рассматриваться не как единый краткий момент истории, а как длительная и сложная цепь событий. Американский исследователь Д. Мельтцер (Meltzer, 1989) определил процесс постепенного проникновения в Новый Свет в разное время небольших групп прото-америндов как «migratory dribbles», что можно приблизительно передать на русском языке как «миграционные капли» или «капельная миграция». Такая модель, как мне представляется, наиболее точно отражает картину формирования антропологического и генетического своеобразия аборигенного населения Америки, включая большой размах межгрупповой изменчивости. Я склонен согласиться с мнением X. Комаса, что никогда «не было и нет американского гомотипа». Об этом свидетельствует разнообразие индейских групп почти по всем системам признаков. Приведу еще один пример. По «классическим» нормам, выведенным на основании данных А. Грдлички, высота лица и скуловой диаметр на черепах америндов должны укладываться в рамки 69–77 мм и 139–145 мм соответственно. Однако, те же размеры в большой серии черепов древних перуанцев оказались равными 64,4 и 132,0 мм (Burger & Ziegelmayer, 1982). Конечно, могут объяснить такие явления локальными тенденциями микроэволюции, но здесь речь идет уже не просто о колебаниях размерных показателей, но о существенных различиях, имеющих таксономически значительный масштаб.
Разнообразие индейского населения Америки по данным антропологии и генетики не может быть свдено к микроэволюционным процессам в пределах Старого Света и имеет глубокие корни в исходных характеристиках древних мигрантов, о чем сейчас убедительно свидетельствуют работы бразильских антропологов. Особенно это относится к различиям аборигенного населения Северной и Южной Америки. Если генетикам удалось обнаружить хорошо выраженное сходство индейцев Северной Америки с народами восточной Сибири, то исследования митохондриальной ДНК у индейцев Южной Америки выявляет параллели с тихоокеанскими популяциями, что показано в работах французского генетика Э. Денамюра (Denamur, 1994). Исследования Американско-Индийской экспедиции на Новой Гвинее и в Южной Америке установили генетическое сходство между папуасами и индейцами Южной Америки, что, по мнению участников этого проекта, является показателем общего происхождения обеих групп (Bahatra et al., 1995)
Сложность процесса первоначального заселения Америки, включая разные пути миграций и полиморфизм их участников, отразилась на современных оценках времени «открытия Америки». Приведем для иллюстрации этого положения некоторые мнения археологов и генетиков: 50–40 тыс. лет (Медведев, 1979), 40–16 тыс. лет (Williams et al., 1985), 34–17 тыс. лет (Schurret al., 1993), 27 тыс. лет (Szathmary, 1981), не ранее 40 тыс. лет (Stewart, 1981), 20 тыс. лет (Сукерник, Кроуфорд), 14–12 тыс. лет (Turner, 1981; Dixon, 1993). Если учесть наличие двух возможных типов инвазии в Новый Свет (кратковременное посещение и колонизация), то большинство цитируемых здесь исследователей окажутся правыми. Древнейшие малочисленные группы первопроходцев, часть которых, была занесена на Американский берег случайно, вернее всего вымирали, т. к. их материальная культура и уровень адаптабельности не позволяли долго продержаться и выжить в условиях нового мира, но все же именно их мы должны считать «первооткрывателями Америки». Вероятная дата их прибытия в Новый Свет, по моему мнению, — 30–35 тыс. лет, в эпоху становления на Земле «поведенчески современного человека», когда были «открыты» и заселены многие острова в Тихом океане и стали развиваться древнейшие средства навигации. По-видимому, первая высадка людей на землю Нового Света состоялась в тропиках и, возможно, была связана с Межпассатным экваториальным противотечением. Путь этот был трудным, но, как справедливо замечает Дж. Диксон, северный вариант — через Берингию — в те времена был еще труднее. Сейчас невозможно решить, какой континент был «открыт» первым — Северная или Южная Америка. Думаю, что скорее всего, это была Центральная Америка, куда могло занести людей Межпассатное теплое течение. И к северу и к югу мореплавателям помешали бы холодные океанические течения, в частности направить свои еще очень примитивные суда сразу к южноамериканскому берегу им воспрепятствовало бы мощное холодное течение Гумбольдта. Повернув на север, они столкнулись бы примерно с такого же рода препятствием. Вероятно, верхнепалеолитические мореходы предпочли высадиться там, где увидели землю, тем более, что они были утомлены тяжелым путешествием и торопились пополнить запасы воды и пищи. Далее движение группы (и последующих групп) могло происходить по суше как в северном, так и в южном направлениях, о чем мы уже можем судить по конкретным археологическим и антропологическим находкам в Южной и Северной Америке. К сожалению, эта первая смелая попытка заселения Нового Света оказалась безуспешной. Однако, можно предположить, что «капельные миграции» с временными эпизодами пребывания в Америке продолжались и после первого путешествия. Более многочисленные волны хорошо подготовленных переселенцев хлынули в Новый Свет только по прошествии ряда тысячелетий разными путями, в разное время. Некоторым из них удалось внести свой вклад в становление сложного, разнородного генофонда коренного населения Америки.
1. Дискуссия о времени и месте проникновения первых людей в Америку, несмотря на значительные достижения в области антропологии, генетики и археологии, пока далека от завершения. Сформулированная и детально разработанная в основном археологами модель инвазии человека в Новый Свет в голоцене — 12–14 тыс. лет назад через область Берингии отражает лишь определенный этап межконтинентальной дисперсии человечества, но не объясняет многих фактов, относящихся к истории формирования аборигенного населения Америки.
2. Выдвинутая в прошлом веке гипотеза «американского гомотипа», согласно которой все индейское население гомогенно в отношении антропологического типа и может быть отнесено к монголоидной расе, не выдержала испытание временем: исследования в области краниологии, генетики, одонтологии, дерматоглифики обнаружили очень высокий уровень разнообразия аборигенного населения Нового Света, что трудно объяснить одними лишь микроэволюционными процессами в пределах этой части Света.
3. В науке существовало и существует несколько различных точек зрения относительно места проникновения первых людей в Америку: из Северо-Восточной Азии через Берингию, из Европы через Гренландию, из Австралии через Антарктиду или через Тихий океан. Приоритетным из этих предположений пока является Берингоморская гипотеза, принятая исследователями в США и Европе и по смыслу связанная с идеей «американского гомотипа». В Латинской Америке, кроме того, остается достаточно популярной гипотеза французского исследователя Поля Риве, согласно которой помимо берингоморского пути расселения, имели место миграции со стороны Австралии, Меланезии и Полинезии. Эта точка зрения, учитывающая многообразие антропологических типов мигрантов в Америку, имеет достаточно серьезную аргументацию, но долгое время не была принята в науке. В настоящее время становится общепризнанным фактом наличие австрало-меланезийского компонента в составе коренного населения Америки (особенно Южной). В этом вопросе гипотезу П. Риве поддерживают современные бразильские антропологи, хотя они скорее склоняются к модели проникновения экваториальных элементов через Берингию во время раннего этапа расселения. Некоторые антропологи придерживаются сходной позиции, но считают, что монголоидные и австрало-меланензийокие элементы проникли в Америку через Берингию в составе одной миграции, являясь носителями недифференцированного австрало-монголоидного комплекса, распространенного в конце плейстоцена в Юго-Вострчной и Восточной Азии. Бразильские коллеги предпочитают гипотезу инвазии обоих компонентов в составе двух последовательных независимых волн. Наличие исходного недифференцированного комплекса в Азий подтверждается антропологическим материалом (хорошо сохранившиеся черепа из Верхнего грота пещеры Чжоукоудянь). Некоторые археологи и антропологи, базируясь на современных данных археологии и генетики, не отвергают также предположения П. Риве о возможности морской миграции через Тихий океан из Австралии, Новой Гвинеи и с океанийских островов.
4. Продолжаются споры о числе волн мигрантов, заселявших Америку. Популярна «трехволновая» гипотеза: первая волна — основное индейское население; вторая — предки индейцев языковой семьи на-дене; третья волна — Предки эскимосов и алеутов. Некоторые исследователи полагают, что необходимо отдельно рассматривать палеоамериканскую волну, с которой в Америку проник экваториальный компонент. Деление на волны, разумеется, довольно условно. Антропологи и археологи все больше прислушиваются к мнению сторонников «гипотезы капельных миграций» американского исследователя Д. Мельтцера, предположившего, что последовательных миграций, причем разных по антропологическому составу, моглс быть много, и весь процесс переселения занял большой отрезок времени.
5. Среди современных специалистов нет также согласия относительно времени первоначального заселения Америки. Диапазон точек зрения здесь весьма широк — от 40 (или даже 50 тыс.) до 14–12 тыс. лет назад. Последняя дата принята у археологов, но она все же не учитывает датировок ряда древнейших находок в Северной и Южной Америке. Вероятно, наиболее приемлемой датой можно было бы считать 30–35 тыс. лет до современности, т. к. на этот период приходится пик миграционной активности верхнепалеолитических популяций и, по-видимому, приобретение человеком первых навыков мореплавания, о чем мы можем судить по времени заселения островов в районе Восточной Азии и Западной Полинезии (см. гл. 3).
6. Первые этапы освоения человеком Америки, вероятно, представляли собой многочисленные безуспешные попытки колонизации новых территорий, причем прибывшие в этот регион малочисленные группы мигрантов, по-видимому, большей частью вымирали. Вследствие этого современная наука располагает весьма скудными материалами того периода и не может установить преемственность между древнейшими и более поздними инвазиями.
Алексеев В. П. География человеческих рас. М., 1974
Бунак В. В. Череп человека и стадии его формирования у ископаемых людей и современных рас. М., 1959
Бунак В. В., Герасимова М. М. Верхнепалеолитический череп Сунгирь 1 и его место в ряду других верхнепалеолитических черепов. // Сунгирь. Антропологическое исследование. М. 1984. С. 14–100.
Дебец Г. Ф. Происхождение коренного населения Америки. // Народы мира. Народы Америки, том 1. М., 1959. С. 9–24.
Деникер И. Человеческие расы (перевод с французского). СПб. 1902.
Зубов А. А. О физическом типе древнейшего населения Америки. // Сов. этнография. № 4, 1968.
Зубов А. А. Одонтологические данные по вопросу о двух первичных очагах расообразования. // Ранняя этническая история народов Восточной Азии. М., 1977. С. 69–73.
Зубов А. А. «Восточный зубной комплекс» у населения Тихоокеанского побережья Перу. // Тезисы доклада на XIV Тихоокеанском Конгрессе. М., 1979. С. 194–195.
Зубов А. А., Халдеева Н. И. Одонтология в современной антропологии. М., 1989.
Зубов А. А., Халдеева Н. И. Одонтология в антропофенетике. М., 1993.
Зубов А. А. Древнейшее заселение Америки в свете данных физической антропологии. // Российский этнограф. Вып. 20. 1993.С. 187–202.
Зубов А. А. Проблема термина «раса» и расовых классификаций в современной физической антропологии. // Этнографическое обозрение. 1996. № 1. С. 15–24.
Зубов А. А. Биолого-антропологическая характеристика коренного до-европейского населения Америки. // Население Нового Света: проблемы формирования и социо-культурного развития. М., 1999. С. 11-66.
Зубов А. А. Некоторые спорные моменты в традиционных взглядах на формирование физического типа американских индейцев. // История и семиотика индейских культур Америки. М., 2002. С. 388–399.
Зубов А. А. Древность Homo sapiens в Азии. // Вестник антропологии. 2005. Вып. 12. С. 12–22
Медведев Г. И. О времени и степени возможного участия палеолитических обитателей Сибирского плоскогорья в происхождении древних культур Северной Америки. // тезисы доклада на XIV Тихоокеанском Конгрессе. М., 1979. С. 202–204.
Пласкетт Д., Диксон Д. Люди вне Азии и раннее заселение Америки. // Тезисы доклада на XIV Тихоокеанском Конгрессе. М., 1979. С. 212.
Рогинский Я. Я. Проблема происхождения монгольского расового типа. // Антропологический журнал. № 2. 1937
Рогинский Я. Я., Левин М. Г. Антропология. М., 1978.
Спицын В. А. Биохимический полиморфизм человека. М., 1985.
Сукерник Р. И., Кроуфорд М., Осипова Л. П., Вибе Л. П., Шенфилд М. С. Первоначальное заселение Америки в свете данных популяционной генетики. // Экология американских индейцев и эскимосов. Проблемы индеанистики. М., 1988.
Хейердал Т. Аку-Аку. М., 1959.
Хить Г. Л., Долинова Н.А. Расовая дифференциация человечества по данным дерматоглифики. М. 1990.
Bahatra К., Black F. L., Smith Th. A., Prasad M. L., Koki G. N. Class I HLA in two long separated populations: Melanesians and South Amerinds. // American Journal of Physical Anthropology. 1995. Vol. 97. N 3. P. 291–305
Bianchi N.O., Bailliet G., Bravi C.M. Peopling of the Americas as inferred through the analysis of the mitochondrial DNA // Revista bras, genetic.1995. Vol. 18. N 4. P. 661–668.
Birdsell J. B. The problem of the early peopling of the Americas as viewed from Asia. N. Y., 1951.
McBrearty S. & Brooks A. S. The revolution that wasn’t: a new interpretation of the origin of modern human behaviour // / Journal of Human Evolution. 2000. Vol. 39. P. 453–563.
Brothwell D., Burleigh R. On sinking Otovalo man. // Journal of Archaeological Sciences. 1977. Vol. 4. N 3. P. 291–294.
Bryan A. L. An overview of palaeoamerican prehistory from a circumpa-cific spectrum. // Early man in America. Alberta. 1978.
Burger E., Ziegelmayer G. Eine umfangreiche Skelettserie aus verschie-denen Kustenregionen Altperus // Homo. 1982. Bd. 33. N 4. P. 220–230.
Castillo H. L. de. Odontometría у morfología dental de los Guajiros. Caracas, 1973.
Christensen A. F. Odontometric microevolution in the Valley of Oaxaca, Mexico. // Journal of Human Evolution. 1998. Vol. 34. P. 333–360.
Comas J. Manual de antropología fisíca. Mexico. 1966.
Cunningham D. L., Jantz R. L. The morphometric relationship of Upper Cave 101 and 103 to modern Homo sapiens. // Journal of Human Evolution. 2003. Vol. 45. P. 1–18.
Davies D. M. Some observations on the Otovalo skeleton and other remains in South America. // Journal of Human Biology. 1978. Vol. 7. N 4. P. 279–281.
Denamur E. Les gènèticiens sur la piste des premiers Americains // Journal of medical sciences. 1994. Vol. 10. N 11. P. 1157–1160.
Devoto F. C. H., Arias N. H., Ringuelet S., Palma N. H. Shovel-shaped incisors in a Northwestern Argentina population. // Journal of Dental Re-seach. 1968. N 5.
Diaz Ungria A. G. de. Estudio comparativo de las características serologicas у morfologic as correspondientes a las poblaciones Guajiro, Guahibo, Guarao у Yaruro. // Universidad Central de Venezuela. Caracas, 1966.
Dillehay T. D., Collins M. B. Early cultural evidence from Monte Verde in Chile. // Nature. 1988. Vol. 332. N 6160. 150–152.
Dixon E. J. Quest for the origins of the first Americans. Albuquerque. University of New Mexico, 1993.
Escobar V., Conneally М., Lopez C. The dentition of the Queckchi indians. Anthropological aspects. // American Journal of Physical Anthropology. 1977. Vol. 47. N 3. P. 443–451.
Gam S. Human races. Springfield, 1962.
Garruto R. M., Plato C. C., Hoff C., Newman M.T., Gajdusek D. C., Baker P. T. Characterization and distribution of dermatoglyphic features in Eskimo, North, Central and South American Indians. // Dermatoglyphics — Fifty years later. N. Y. 1979. Vol. 15. N 6.
Guèrin C. Les plus anciens Americains et leur environnement. // Bull. Mensuelle Soc. Anthropol. 1993. Vol. 62. N 8. P. 275–278.
Gusinde M. Ein Feuerland-Schädel neu interpretiert. // Ann. Naturhist. Mus. Wien. 1975. Bd. 70. P. 665–667.
Hooton E. A. The Indians of Pecos Pueblo. A study of their skeletal remains. New Haven. 1930.
Howells W. W. The origin of American Indian racial types. The Maya and their neighbours. N. Y. 1940.
Hrdlička A. Shovel-shaped teeth. // American Journal of Physical Anthropology. 1920. Vol. 3. P. 429–465.
Imbelloni J. Tabla clasificatoria de los Indios. Regiones biológicas у grupos raciales humanos en America // Physis. 1938. Vol. 12. P. 229–249. Buenos Aires.
Imbelloni J. The peopling of America. // Acta Americana. 1943. Vol. l. P. 309–330.
Irving R. Peopling of the Americas // Quaternary Reseach. 1976. Vol. 6. N 4. P. 597–612.
Jacob T. Racial identification of the Bronze Age human dentitions from Bali, Indonesia. // Journal of Dental Research. 1967. Vol. 46. N 5. Part 1. P. 66–79.
Kieser J. A., Preston С. B. The dentition of the Lengua Indians of Paraguay. // American Journal of Physical Anthropology. 1981. Vol. 55. N 4. P. 485–490.
Lahr M. Patterns of modern human diversification. Implication for Amerindian origins. // Yearbook of Physical Anthropology. 1995. Vol. 38. Suppl. N 21. P. 163–198.
Mansalve М. V., Groot de Restrepo H., Espinel A., Correal G., Devine D. V. Evidence of mitochondrial DNA diversity in South American aboriginals. // Annals of Human Genetics. 1994. Vol. 58. N 3. P. 265–273.
Meltzer D. J. Why don’t we know when the first people came to North America? // American Antiquity. 1989. Vol. 54.
Mendes Correa A. A. Nouvelle hypothese sur le peuplement de l’Amerique du Sud // Annales Facult. Ciences de Porto. 1928. Vol. 15. P. 5–31.
Molnar S. Human variation. Races, Types, and Ethnic Groups. Prentice Hall. New Jersey. 1992.
Neel J. V. Diversity within and between South American Indian tribes. // Journal of Medical Sciences. 1973. Vol. 9. N 9-10. P. 1216–1224.
Nei M. & Roychoudhury A.K. Genetic relationship and evolution of human races. // Evolutionary Biology. 1982. Vol. 4.
Neves W. A., Pucciarelli H.M. Extra-continental biological relationships of early South American human remains. A multivariate analysis. // Ciencia e Cultura. 1989. Vol. 41. P. 566–575.
Neves W. A., Meyer D., Pucciarelli H. M. The contribution of the morphology of early South and North American skeletal remains to the understanding of the peopling of the Americas // American Journal of Physical Anthropology. 1993. Suppl. N 16. P. 150–151
Neves W. A., Powell J. F., Ozolins E. G. Lapa Vermelha IV Hominid I: Morphological affinities of the earliest known American // American Journal of Physical Anthropology. 1998. Suppl. 26. P. 169.
Neves W. A., Prous A., Gonzàlez-José R., Kipnis R., Powell J. Early Ho-locene human skeletal remains from Santana do Riacho, Brazil: implications for the settlement of the New World // / Journal of Human Evolution. 2003. Vol. 45. P. 19–42.
Neves W. A., Hubbe М., М. M. Okumura, Gonzalez-Jose R., Figuti L., Eggers S., Dantas de Blasis P. A. A new early Holocene human skeleton from Brazil: implications for the settlement of the New World. // Journal of Human Evolution. 2005. Vol. 48. P. 403–414.
Neves W. A., Hubbe M., Beethoven Piló L. Early Holocene human skeletal remains from Sumidouro Cave, Lagoa Santa, Brazil: History of discoveries, geological and chronological context, and comparative cranial morphology. // Journal of Human Evolution. 2007. Vol. 52. P. 16–30.
Newman M. T. The application of ecological rules to the anthropology of the aboriginal New World. // American Anthropologist. 1951. Vol. 55. P. 201–212.
Pereira da Silva M.A. Les dermatoglyphes digito-palmares des Indians Alakaluf des archipels de Patagonie occidentale. Bull, et mem. Soc. anthropol. Paris. 1974. Vol. 1. N 1. 85-108.
Plhak M. On the morphology of the fossil hominids of Laguna Beach and Los Angeles. // Journal of Human Evolution. 1980. Vol. 9. N 8. P. 585–589.
Pucciarelli H.M., Gonzalez-Jose R., Neves W. A., Sardi M.L., Rozzi F.R. East-West cranial differentiation in pre-Columbian populations from Central and North America. // Journal of Human Evolution. 2008. Vol. 54. P. 296–308.
Quillici J. C. Hemotypologie des populations andines et du Piemont andin. Paris, 1977.
Rivet P. Los origenes del hombre americano. Mexico. 1978.
Rodriguez J. V. C. Algunos aspectos metodologicos-bioantropologicos relacionados con el poblamiento de America. // Maguare. Revista del De-partamento de Antropologia. Universidad Nacional de Colombia. 1987. N 5. P. 11–17.
Rogers S. L. The Yuha human fossil from Southern California. // Measurements and relationships. 1977. Vol. 47. N 1. P. 157.
Rothhammer F., Lassere E., Blanco R., Covarrubias E., Dixon AL. Microevolution in human Chilean populations. Shovel shape, Mesial palatal version and other dental traits in Pewenche Indians. // Zeitschrift fur morphologie und Anthropologie. 1968. Bd. 60. N 2.
Schurr T. G., Torroni A., Cabell ALA, Brown M.D., Wall-Lace D.C. Mitichondrial DNA variation and the human colonization of Siberia and the Americas. // American Journal of Physical Anthropology. 1993. Suppl. N 6. P. 127.
Szathmary E. J. E. Genetic markers in Siberian and North American populations. // Yearbook of Physical Anthropology. 1981. N 24.
Stewart T. D. Evolutionary status of the first Americans // American Journal of Physical Anthropology. 1981. Vol. 56. N 4. P. 461–466.
Turner Ch.G.II. Dental evidence for the peopling of Americas. // Early man in the New World. San Diego, 1981.
Valle ALAL Human life zones. Lima, Perw, 1964.
Williams R. C., Steinberg A. G., Gershovtz H., Bennet P. H., Knowler W. C., Pettitt D. J., Butler W., Baird R., Dowda-Rea L., Burch Th. A., Morse G., Smith Ch. G. Gm Allotypes in native Americans. Evidence for three distinct migrations across the Bering Land bridge. // American Journal of Physical Anthropology. 1985. Vol. 66. N 1. P. 1–19.
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги