Глава 25. Механизмы видообразования
В предыдущей главе было описано, каким образом Дарвин пришел к выводу о существовании у растений и животных наследственной изменчивости как при искусственном разведении, так и в природных популяциях. Он понимал, что наследственные изменения должны играть важную роль в эволюции, но не мог предложить механизм, который объяснял бы их возникновение при сохранении дискретности признаков. Лишь после того, как были вторично открыты работы Менделя о наследственности и оценено их значение для эволюционной теории, появилась возможность разрешить многие из этих проблем. Современное объяснение изменчивости живых организмов — это результат синтеза эволюционной теории, основанной на работах Дарвина и Уоллеса, и теории наследственности, основанной на законах Менделя. Сущность изменчивости, наследственности и эволюции можно теперь объяснить с помощью данных, полученных в одной из областей биологии, известной под названием популяционной генетики.
25.1. Популяционная генетика
Популяция — это группа организмов, принадлежащих к одному и тому же виду и занимающих обычно четко ограниченную географическую об-ласть. Дарвина интересовало, каким образом естественный отбор, действуя на уровне отдельного организма, вызывает эволюционное изменение. После вторичного открытия работ Менделя, доказавших корпускулярную природу наследственности, большое внимание при изучении изменчивости, наследственности и эволюционных изменений стали уделять генотипу. Бэтсон, который в 1905 г. ввел термин "генетика", видел задачу этой науки в "освещении явлений наследственности и изменчивости".
Основу современной эволюционной теории, которую называют неодарвинизмом или синтетической теорией эволюции, составляет изучение популяционной генетики. Гены, действуя независимо или совместно с факторами среды, определяют фенотипические признаки организмов и обусловливают изменчивость в популяциях. Фенотипы, приспособленные к условиям данной среды или "экологическим рамкам", сохраняются отбором, тогда как неадаптивные фенотипы подавляются и в конце концов элиминируются. Естественный отбор, влияя на выживание отдельных особей с данным фенотипом, тем самым определяет судьбу их генотипа, однако лишь общая генетическая реакция всей популяции определяет выживание данного вида, а также образование новых видов. Только те организмы, которые, прежде чем погибнуть, успешно произвели потомство, вносят вклад в будущее своего вида. Для истории данного вида судьба отдельного организма не имеет существенного значения.
25.1.1. Генофонд
Генофонд слагается из всего разнообразия генов и аллелей, имеющихся в популяции, размножающейся половым путем; в каждой данной популяции состав генофонда из поколения в поколение может постоянно изменяться. Новые сочетания генов образуют уникальные генотипы, которые в своем физическом выражении, т. е. в форме фенотипов, подвергаются давлению факторов среды, производящим непрерывный отбор и определяющим, какие гены будут переданы следующему поколению.
Популяция, генофонд которой непрерывно изменяется из поколения в поколение, претерпевает эволюционное изменение. Статичный генофонд отражает отсутствие генетической изменчивости среди особей данного вида и отсутствие эволюционного изменения.
25.1.2. Частоты аллелей
Любой физический признак, например окраска шерсти у мышей, определяется одним или несколькими генами. Каждый ген может существовать в нескольких различных формах, которые называют аллелями (см. табл. 23.2). Число организмов в данной популяции, несущих определенный аллель, определяет частоту данного аллеля (которую иногда называют частотой гена, что менее точно). Например, у человека частота доминантного аллеля, определяющего нормальную пигментацию кожи, волос и глаз, равна 99%. Рецессивный аллель, детерминирующий отсутствие пигментации — так называемый альбинизм, — встречается с частотой 1%. В популяционной генетике частоту аллелей или генов чаще выражают не в процентах или простых дробях, а в десятичных дробях. Таким образом, в данном случае частота доминантного аллеля равна 0,99, а частота рецессивного аллеля альбинизма — 0,01. Общая частота аллелей в популяции составляет 100%, или 1,0, поэтому

Как это принято в классической генетике, аллели можно обозначить буквами, например доминантный аллель (нормальная пигментация) — буквой N, а рецессивный (альбинизм) — буквой n. Для приведенного выше примера частота N = 0,99, а частота n = 0,01.
Популяционная генетика заимствовала у математической теории вероятностей два символа, р и q, для выражения частоты, с которой два аллеля, доминантный и рецессивный, встречаются в генофонде данной популяции. Таким образом,
p + q= 1,
где:
р-частота доминантного, а q-частота рецессивного аллеля.
В примере с пигментацией у человека р = 0,99, а q = 0,01;
Р + q = 1
0,99 + 0,01 = 1
Значение этого уравнения состоит в том, что, зная частоту одного из аллелей, можно определить частоту другого. Пусть, например, частота рецессивного аллеля равна 25%, или 0,25. Тогда
Р + q = 1
р + 0,25 = 1
р = 1-0,25
р = 0,75
Таким образом, частота доминантного аллеля равна 0,75, или 75%.
25.1.3. Частота генотипов
Частоты отдельных аллелей в генофонде позволяют вычислять генетические изменения в данной популяции и определять частоту генотипов. Поскольку генотип данного организма — главный фактор, определяющий его фенотип, вычисление частоты генотипа используют для предсказания возможных результатов тех или иных скрещиваний. Это имеет важное практическое значение в сельском хозяйстве и медицине.
Математическая зависимость между частотами аллелей и генотипов в популяциях была установлена в 1908 г. независимо друг от друга английским математиком Дж. Харди и немецким врачом В. Вайнбергом. Эту зависимость, известную под названием равновесия Харди Вайнберга, можно сформулировать так: частоты доминантного и рецессивного аллелей в данной популяции будут оставаться постоянными из поколения в поколение при наличии определенных условий. Условия эти следующие:
1) размеры популяции велики;
2) спаривание происходит случайным образом;
3) новых мутаций не возникает;
4) все генотипы одинаково плодовиты, т.е. отбора не происходит;
5) поколения не перекрываются;
6) не происходит ни эмиграции, ни иммиграции, т. е. отсутствует обмен генами с другими популяциями.
Поэтому любые изменения частоты аллелей должны быть обусловлены нарушением одного или нескольких из перечисленных выше условий. Все эти нарушения способны вызвать эволюционное изменение; и если такие изменения происходят, то изучать их и измерять их скорость можно с помощью уравнения Харди-Вейнберга.
25.1.4. Уравнение Харди-Вайнберга
Это уравнение дает простую математическую модель, которая объясняет, каким образом в генофонде сохраняется генетическое равновесие; но главное применение его в популяционной генетике — вычисление частот аллелей и генотипов.
Если имеется два организма, один гомозиготный по доминантному аллелю А, а другой — по рецессивному аллелю а, то все их потомки будут гетерозиготными (Аа):

Если наличие доминантного аллеля А обозначить символом р, а рецессивного аллеля а — символом q, то картину скрещивания между особями F1, возникающие при этом генотипы и их частоты можно представить следующим образом:

Поскольку аллель А доминантный, отношение доминантных генотипов к рецессивным составляет 3:1 — это менделевское отношение при моногибридном скрещивании. Используя символы р и q, результаты приведенного выше скрещивания можно представить следующим образом:
р2 — доминантные гомозиготы;
2pq — гетерозиготы;
q2 — рецессивные гомозиготы.
Такое распределение возможных генотипов носит статистический характер и основано на вероятностях. Три возможных генотипа, образующихся при таком скрещивании, представлены со следующими частотами:

Сумма частот трех генотипов, представленных в рассматриваемой популяции, равна единице; пользуясь символами р и q, можно сказать, что вероятности генотипов следующие:
p2 + 2pq + q2 = 1,
На математическом языке р + q = 1 представляет собой уравнение вероятности, тогда как р2 + 2pq + + q2 = 1 является квадратом этого уравнения [т. е. (р + q)2].
Поскольку
p — частота доминантного аллеля;
q — частота рецессивного аллеля;
p2 — гомозиготный доминантный генотип;
2pq — гетерозиготный генотип;
q2 — гомозиготный рецессивный генотип,
можно вычислить частоты всех аллелей и генотипов, пользуясь выражениями
для частот аллелей: р + q = 1;
для частот генотипов: р2 + 2pq + q2 = 1.
Однако для большинства популяций частоту обоих аллелей можно вычислить только по доле особей, гомозиготных по рецессивному аллелю, так как это единственный генотип, который можно распознать непосредственно по его фенотипическому выражению.
Например, один человек из 10 000 альбинос, т.е. частота альбинотического генотипа составляет 1 на 10 000. Поскольку аллель альбинизма рецессивен, альбинос должен быть гомозиготным по рецессивному гену, т.е. на языке теории вероятности

Зная, что q2 = 0,0001, можно определить частоты аллеля альбинизма (q), доминантного аллеля нормальной пигментации (p), гомозиготного доминантного генотипа (р2) и гетерозиготного генотипа (2pq). Так как

т.е. частота аллеля альбинизма в популяции равна 0,01 или 1%. Поскольку

частота доминантного аллеля в популяции равна 0,99, или 99%. А если

т.е. частота гетерозиготного генотипа составляет 0,0198; иными словами, примерно 2% индивидуумов в данной популяции несут аллель альбинизма либо в гетерозиготном, либо в гомозиготном состоянии.
Как показывают эти вычисления, частота рецессивного аллеля в популяции неожиданно велика при малом числе индивидуумов с гомозиготным рецессивным генотипом.
Гетерозиготных индивидуумов, нормальных по фенотипу, но обладающих рецессивным геном, который в гомозиготном состоянии может вызвать нарушение метаболизма, называют носителями. Как показывают вычисления с использованием уравнения Харди-Вайнберга, частота носителей в популяции всегда выше, чем можно было бы ожидать на основании оценок частоты фенотипического проявления данного дефекта. Это ясно видно из табл. 25.1.

Таблица 25.1. Некоторые наследственные метаболические дефекты и частоты рецессивных гомозиготных и гетерозиготных генотипов
25.1. Кистозный фиброз поджелудочной железы встречается среди населения с частотой 1 на 2000. Вычислите частоту носителей.
25.1.5. Следствия уравнения Харди-Вайнберга
Из уравнения Харди-Вайнберга следует, что значительная доля имеющихся в популяции рецессивных аллелей находится у гетерозиготных носителей. Фактически гетерозиготные генотипы служат важным потенциальным источником генетической изменчивости. Это приводит к тому, что в каждом поколении из популяции может элиминироваться лишь очень малая доля рецессивных аллелей. Только те рецессивные аллели, которые находятся в гомозиготном состоянии, проявятся в фенотипе и тем самым подвергнутся селективному воздействию факторов среды и могут быть элиминированы. Многие рецессивные аллели элиминируются потому, что они неблагоприятны для фенотипа — обусловливают либо гибель организма еще до того, как он успеет оставить потомство, либо "генетическую смерть", т.е. неспособность к размножению.
Однако не все рецессивные аллели неблагоприятны для популяции. Например, у человека из всех групп крови чаще всего встречается группа О, соответствующая гомозиготности по рецессивному аллелю. Другим примером служит серповидноклеточная анемия. Это наследственное заболевание крови, широко распространенное в ряде областей Африки и Индии, в некоторых средиземноморских странах и у негритянского населения Северной Америки. Индивидуумы, гомозиготные по соответствующему рецессивному аллелю, обычно умирают, не достигнув половой зрелости и элиминируя таким образом из популяции по два рецессивных аллеля. Что касается гетерозигот, то они не гибнут. Установлено, что во многих частях земного шара частота аллеля серповидноклеточности остается относительно стабильной. У некоторых африканских племен частота гетерозиготного фенотипа достигает 40%. Раньше думали, что этот уровень поддерживается за счет появления новых мутантов. Однако в результате дальнейших исследований выяснилось, что дело обстоит иначе: оказалось, что во многих частях Африки, где среди факторов, угрожающих здоровью и жизни, важное место занимает малярия, люди, несущие аллель серповидноклеточности, обладают повышенной резистентностью к этой болезни. В малярийных районах Центральной Америки это селективное преимущество гетерозиготного генотипа поддерживает частоту аллеля серповидноклеточности среди населения на уровне 10-20%. У североамериканских негров, которые уже 200-300 лет не испытывают на себе селективного эффекта малярии, частота аллеля серповидноклеточности упала до 5%. Это снижение можно частично отнести на счет обмена генами в результате браков между представителями черной и белой расы, однако важным фактором служит отсутствие в Северной Америке малярии, устраняющее селективное давление в пользу гетерозигот; в результате рецессивный аллель медленно элиминируется из популяции.
Этот пример эволюции в действии ясно демонстрирует селективное влияние среды на частоту аллелей — механизм, нарушающий генетическое равновесие, предсказываемое законом Харди-Вейнберга. Именно такого рода механизмы вызывают в популяциях сдвиги, ведущие к эволюционному изменению.
25.2. Факторы, вызывающие изменения в популяциях
Принцип равновесия Харди-Вайнберга гласит, что при наличии определенных условий частота аллелей остается постоянной из поколения в поколение. При этих условиях популяция будет находиться в состоянии генетического равновесия и никаких эволюционных изменений происходить не будет. Однако принцип Харди-Вайнберга носит чисто теоретический характер. Очень немногие популяции находятся в условиях, при которых сохраняется равновесие (см. разд. 25.1.3).
В разд. 23.8.4 были подробно описаны четыре главных источника генетической изменчивости: кроссинговер во время мейоза, независимое распределение хромосом при мейозе, случайное оплодотворение и мутационный процесс. Первые три источника часто объединяют под общим названием половой рекомбинации; они обусловливают перетасовку генов, лежащую в основе происходящих изо дня в день непрерывных изменений. Но хотя эти процессы и приводят к образованию новых генотипов и изменяют частоты генотипов, они не вызывают никакого изменения имеющихся аллелей, так что частоты аллелей в популяции остаются постоянными.
Многие эволюционные изменения, однако, происходят вслед за появлением новых аллелей, а главным источником последних служат мутации.
Условия, необходимые для равновесия Харди-Вайнберга, нарушаются и в ряде других случаев: когда скрещивание носит неслучайный характер; когда популяция мала, что ведет к дрейфу генов; когда генотипы обладают различной фертильностью, что создает генетический груз; при наличии обмена генами между популяциями. Ниже рассматривается каждая из этих ситуаций.
25.2.1. Неслучайное скрещивание
В большинстве природных популяций спаривание происходит неслучайным образом. Во все тех случаях, когда наличие одного или нескольких наследуемых признаков повышает вероятность успешного оплодотворения гамет, имеет место половой отбор. У растений и животных существует много структурных и поведенческих механизмов, исключающих чисто случайный подбор родительских особей. Например, цветки, у которых лепестки крупнее и нектара больше, чем обычно, вероятно, будут привлекать больше насекомых, что повысит вероятность опыления и оплодотворения. Характер окраски насекомых, рыб и птиц и особенности их поведения, связанные с постройкой гнезда, охраной территории и брачными церемониями, повышают избирательность при скрещивании.
Влияние неслучайного скрещивания на генотип и на частоту аллелей демонстрируют, например, эксперименты, проведенные на дрозофиле. В культуре мух, содержавшей вначале равное число красноглазых и белоглазых самцов и самок, через 25 поколений исчезли все белоглазые особи. Как показали наблюдения, и красноглазые, и белоглазые самки предпочитали спариваться с красноглазыми самцами. Таким образом, половой отбор как механизм избирательного скрещивания обеспечивает некоторым особям более высокий репродуктивный потенциал, в результате чего вероятность передачи генов этих особей следующему поколению повышается. Репродуктивный потенциал особей с менее благоприятными признаками понижен, и передача их аллелей последующим поколениям происходит реже.
25.2.2. Дрейф генов
О дрейфе генов говорят в тех случаях, когда изменения частоты генов в популяциях бывают случайными и не зависят от естественного отбора. Случайный дрейф генов, или эффект Сьюэлла Райта (названный по имени американского генетика, который понял его роль в эволюции), может служить важным механизмом эволюционных изменений в не-больших или изолированных популяциях. В небольшой популяции могут быть представлены не все аллели, типичные для данного вида. Случайные события, например, преждевременная гибель особи, бывшей единственным обладателем какого-то аллеля, приведут к исчезновению этого аллеля в популяции. Если данный аллель встречается в популяции из 1000000 особей с частотой, скажем, 1% (т.е. q = 0,01), то им будут обладать 10000 особей; а в популяции, состоящей из 100 особей, этот аллель будет иметься только у одной особи, так что вероятность его случайной утраты в малой популяции гораздо выше.
Точно так же, как некий аллель может исчезнуть из популяции, частота его может и повыситься чисто случайным образом. Случайный дрейф генов, как показывает само его название, непредсказуем. Небольшую популяцию он может привести к гибели, а может сделать ее еще более приспособленной к данной среде или усилить ее дивергенцию от родительской популяции. С течением времени возможно образование из нее нового вида под действием естественного отбора. Дрейф генов считают существенным фактором в возникновении новых видов в островных и других репродуктивно изолированных популяциях.
С дрейфом генов связано явление, известное под названием принципа основателя. Оно состоит в том, что при отделении от родительской популяции небольшой ее части последняя может случайно оказаться не вполне типичной по своему аллельному составу. Некоторые аллели в ней могут отсутствовать, а другие будут представлены с непропорционально высокой частотой. Постоянное скрещивание внутри такой пионерной популяции приведет к созданию генофонда, отличающегося по частотам аллелей от генофонда исходной родительской популяции. Дрейф генов обычно снижает генетическую изменчивость в популяции, главным образом в результате утраты тех аллелей, которые встречаются редко. Длительное скрещивание особей внутри малой популяции уменьшает долю гетерозигот и увеличивает долю гомозигот. Примеры действия принципа основателя были выявлены при изучении небольших популяций, образованных в Америке религиозными сектами, эмигрировавшими из Германии в XVIII веке. В некоторых из этих сект браки заключались почти исключительно между членами данной секты. В таких случаях частота ряда аллелей здесь сильно отличается от их частоты среди населения как ФРГ, так и Америки. Например, изученные общины данкеров (религиозная секта, обосновавшаяся в Пенсильвании) состояли примерно из 100 семей каждая; это такие маленькие популяции, что в них должен был происходить дрейф генов. Определение групп крови дало следующие результаты:

Эти данные, по-видимому, отражают результаты дрейфа генов, происходящего в малых популяциях.
Дрейф генов может вести к уменьшению изменчивости в пределах популяции, но он может также увеличить изменчивость в пределах вида в целом. В небольших изолированных популяциях могут возникать нетипичные для основной популяции признаки, которые в случае изменения среды могут дать селективное преимущество. Таким образом, дрейф генов может участвовать в процессе видообразования.
25.2.3. Генетический груз
Существование в популяции неблагоприятных аллелей в составе гетерозиготных генотипов называют генетическим грузом. Как отмечалось в разд. 25.1.5, некоторые рецессивные аллели, вредоносные в гомозиготном состоянии, могут сохраняться в гетерозиготных генотипах и при некоторых условиях среды доставлять селективное преимущество; примером служит аллель серповидноклеточности в местах распространения малярии. Любое повышение частоты рецессивных аллелей в популяции в результате вредных мутаций увеличивает ее генетический груз.
25.2.4. Поток генов
В генофонде скрещивающейся внутри себя популяции происходит непрерывный обмен аллелями между особями. Если частоты аллелей не изменяются в результате мутаций, происходящая при таком обмене перетасовка генов ведет к генетической стабильности или равновесию генофонда. В случае возникновения мутантного аллеля он распростра-нится по всему генофонду в результате случайного оплодотворения.
Такое перемещение аллелей в пределах популяции часто не вполне правильно называют "потоком генов". Строго говоря, этот термин относится к перемещению аллелей из одной популяции в другую в результате скрещивания между членами этих двух популяций. Случайное внесение новых аллелей в популяцию — реципиента и удаление их из популяции-донора изменяет частоту аллелей в обеих популяциях и ведет к повышению генетической изменчивости. Несмотря на то что поток генов вносит в популяции генетическую изменчивость, в смысле эволюционного изменения его действие оказывается консервативным. Распространяя мутантные аллели по всем популяциям, поток генов приводит к тому, что все популяции данного вида приобретают общий генофонд, т.е. различия между популяциями уменьшаются. Поэтому прерывание потока генов между популяциями представляет собой одну из предпосылок для образования нового вида.
Интенсивность обмена генами между двумя популяциями зависит от их пространственной близости и от легкости, с которой организмы или гаметы могут переходить из одной популяции в другую. Например, две популяции могут находиться так близко друг к другу, что скрещивание между ними происходит непрерывно, и тогда в генетическом смысле их можно считать одной популяцией, поскольку они обладают общим генофондом; пример — две популяции улиток, обитающие в соседних садах, разделенных живой изгородью.
Летающим животным и пыльцевым зернам относительно легко активно или пассивно распространяться в новые места. Здесь они могут скрещиваться между собой или с местной популяцией, внося в нее при этом генетическую изменчивость.
25.3. Отбор
Действие отбора можно рассматривать на двух связанных между собой уровнях — на уровне организма и на уровне аллелей.
Отбор — это процесс, благодаря которому организмы, по своей морфологии, физиологии и проведению лучше приспособленные к данной среде, выживают и размножаются а менее приспособленные гибнут или им не удается оставить потомков. Первые передают свои удачные признаки следующему поколению, а вторые — нет. Таким образом, можно видеть, что действие отбора достигается благодаря дифференциальной смертности и дифференциальному размножению. Адаптивное значение отбора состоит в том, что он обеспечивает сохранение тех организмов, которые с наибольшей вероятностью могут обеспечить выживание данного вида, а его эффективность зависит от наличия в популяции фенотипической изменчивости.
При увеличении численности популяции некоторые факторы среды, такие как пища (для животных) или свет (для растений), становятся лимитирующими. Это приводит к конкуренции за ресурсы между членами популяции. Организмы, индивидуальные особенности которых дают им преимущество в конкуренции, будут легче овладевать этими ресурсами, выживут и оставят потомство. Организмы, лишенные таких признаков, окажутся в невыгодном положении и могут погибнуть, прежде чем успеют произвести потомство. Совместное действие лимитирующих факторов среды и численности популяции создает давление отбора, интенсивность которого может быть различной.
Таким образом, отбор — это процесс, определяющий, какие аллели будут переданы следующему поколению благодаря дифференциальным преимуществам, которые они доставляют, экспрессируясь в фенотипах. Давление отбора можно поэтому рассматривать как способ увеличить или уменьшить распространенность какого-либо аллеля в генофонде, и эти изменения частоты аллелей могут вести к эволюционному изменению. Крупные изменения в генотипе возникают в результате распространения в генофонде мутантных аллелей.
Степень отбора и его ход во времени зависят от характера мутантного аллеля и его влияния на данный фенотипический признак. Доминантный аллель сразу проявляется в фенотипе, и тут же начинается его положительный или отрицательный отбор. Если же аллель рецессивен, как это обычно для большинства мутаций, то он не подвергается отбору до тех пор, пока не появится в гомозиготном состоянии; вероятность немедленного появления таких рецессивных гомозигот невелика, и новый аллель может исчезнуть из генофонда, прежде чем они возникнут. Аллель, рецессивный в данных условиях, может сохраниться в популяции и дождаться таких изменений среды, при которых его эффект будет доминантным; если он при этом окажется выгодным для организма, то отбор будет благоприятствовать распространению аллеля в популяции, как, например, в случае серповидноклеточной анемии.
Рецессивный мутантный аллель может быстро распространиться в популяции, если его локус в хромосоме сцеплен (находится в тесной близости) с локусом какого-либо доминантного аллеля, имеющего важное функциональное значение и подвергающегося сильному положительному отбору. При таком сцеплении шансы мутантного аллеля соединиться с другим мутантным аллелем и оказаться в гомозиготном состоянии сильно возрастают (рис. 25.1).

Рис. 25.1. Ускорение распространения в популяции мутантного аллеля (а), сцепленного с доминантным аллелем (В), которому сильно благоприятствует отбор
Фенотипические эффекты мутантных аллелей могут быть различными. Мутации генов, контролирующих важные функции, вероятнее всего, окажутся летальными и будут тотчас же элиминированы из популяции. Эволюционные сдвиги происходят обычно в результате постепенного появления многих мутантных аллелей, производящих небольшие прогрессивные изменения в фенотипических признаках.
Существует три типа отбора: стабилизирующий, направленный и дизруптивный. Смысл этих терминов легче всего пояснить, построив кривые нормального распределения для непрерывной фенотипической изменчивости, наблюдаемой в природных популяциях (рис. 25.2).

Рис. 25.2. Три типа отбора, действующего в популяциях. О — первоначальное положение точки соответствия оптимального фенотипа оптимальным условиям среды; N — новое положение точки соответствия. Особи, признаки которых попадают в заштрихованные участки, оказываются в невыгодном положении и элиминируются отбором. Цифрами 1-3 указана последовательность поколений
25.3.1. Стабилизирующий отбор
Стабилизирующий отбор происходит в тех случаях, когда фенотипические признаки оптимально соответствуют условиям среды и конкуренция относительно слабая. Такой отбор действует во всех попу-ляциях, элиминируя особей с крайними отклонениями признаков. Например, существует некая оптимальная длина крыла для хищной птицы определенных размеров с определенным образом жизни в данной среде. Стабилизирующий отбор, действуя благодаря дифференциальному размножению, будет элиминировать тех птиц, у которых размах крыльев больше или меньше оптимального.
Карн и Пенроуз изучали корреляцию между весом новорожденных и постнатальной смертностью у 13 730 младенцев, родившихся в Лондоне в период 1935-1946 гг. Среди них 614 родились мертвыми или умерли в первый месяц после рождения. Как показывает рис. 25.3, оптимальный для новорожденных вес равен примерно 3,6 кг. Младенцы с большим или меньшим весом оказываются в смысле отбора в невыгодном положении, и смертность среди них несколько повышена. На основании этих данных можно вычислить интенсивность отбора.

Рис. 25.3. Зависимость между весом тела новорожденных и их выживанием. (По M.N. Karn, L.S. Penrose, Ann. Eugen., 1951, 16, 147-164.)
Если в момент рождения или в первый месяц 614 младенцев (из 13 730) умерли, то смертность составляет 4,5%. Даже среди младенцев с оптимальным весом смертность равна 1,8%. Следовательно, ин-тенсивность отбора на вес при рождении, равный 3,6 кг, составляет 4,5% — 1,8% = 2,7%, или 0,027. Среди новорожденных, весящих 1,8 кг, смертность равна 34%, что соответствует интенсивности отбора примерно 30%, или 0,3. Следует указать, однако, что в результате успехов, достигнутых педиатрией после 1946 г., постнатальная смертность значительно снизилась.
Стабилизирующий отбор не способствует эволюционному изменению, а поддерживает фенотипическую стабильность популяции из поколения в поколение.
25.3.2. Направленный отбор
Эта форма отбора возникает в ответ на постепенные изменения условий среды. Направленный отбор влияет на диапазон фенотипов, существующих в данной популяции, и оказывает селективное давление, сдвигающее средний фенотип в ту или другую сторону. После того как средний фенотип придет в оптимальное соответствие с новыми условиями среды, вступает в действие стабилизирующий отбор.
Направленный отбор приводит к эволюционному изменению, оказывая на популяцию такое давление, которое благоприятствует увеличению в ней частоты новых аллелей. Направленный отбор лежит в основе искусственного отбора, при котором избирательное скрещивание особей, обладающих желательными фенотипическими признаками, повышает частоту этих признаков в популяции (см. разд. 25.4). В ряде экспериментов Фальконер выбирал из популяции шестинедельных мышей самых тяжелых особей и давал им спариваться между собой. То же самое он проделывал с самыми легкими мышами. Такое избирательное скрещивание по признаку массы тела привело к созданию двух популяций, в одной из которых масса возрастала, а в другой — уменьшалась (рис. 25.4).

Рис. 25.4. Изменения веса тела в последовательных поколениях в двух популяциях мышей, подвергавшихся отбору по этому признаку. (По D.S. Falconer: J. Genetics, 1953, 51, 470-501.)
После прекращения селекции ни та, ни другая группа не вернулась к первоначальной массе (примерно 22 г). Это показывает, что искусственный отбор по фенотипическим признакам привел к некоторому генотипическому отбору и частичной утрате обеими популяциями каких-то аллелей. В настоящее время непосредственно изучено много примеров классического направленного отбора; они описаны в разд. 25.5.
25.3.3. Дизруптивный отбор
Эта форма отбора, вероятно, встречается реже всего, но она может играть очень важную роль в возникновении эволюционных изменений. Колебания условий среды, связанные, например, со сменой времен года и климатическими факторами, могут благоприятствовать наличию в данной популяции не одного, а двух или нескольких фенотипов. Давле-ние отбора, действующее в самой популяции, может привести к отклонению фенотипов от среднего для данной популяции к обоим крайним типам. В результате популяция окажется разделенной на две субпопуляции. Если поток генов между этими субпопуляциями почему-либо нарушится, каждая из них может дать начало новому виду. В некоторых случаях такая форма отбора приводит к появлению в одной популяции нескольких различных фенотипов — явление, называемое полиморфизмом, которое будет рассмотрено в разд. 25.5.1. В пределах одного вида популяции с различными фенотипами, или экотипы, могут быть адаптированы к определенным условиям среды (см. разд. 25.6.2). Если вид занимает очень обширный географический ареал, у населяющих этот ареал популяций могут наблюдаться локальные изменения фенотипических признаков, в результате чего они становятся промежуточными между популяциями краевых областей.
Такой непрерывный градиент постепенно изменяющихся признаков представляет собой обычно фенотипическую реакцию на изменение климатических и/или эдафических (почвенных) факторов и известен под названием клины (см. разд. 25.6.3).
25.3.4. Интенсивность отбора
Сила давления отбора в данной популяции варьирует в пространстве и во времени, что может быть связано с изменениями внешних или внутренних факторов. К внешним факторам относятся, в частности, увеличение численности хищников или патогенных организмов и конкуренция с другими видами (межвидовая конкуренция) за пищу и места для размножения (у животных) или за свет, воду и минеральные вещества (у растений). Изменения климата или состояния местообитания могут привести к созданию новых давлений отбора. Внутренние факторы, например быстрое повышение численности популяции, нередко приводят к усилению конкуренции за ресурсы (внутривидовая конкуренция). С увеличением популяции данного организма возрастает и численность его паразитов и хищников; кроме того, в многочисленной популяции облегчается передача паразитов и болезней от одной особи к другой. Все эти факторы могут изменять не только силу давления отбора, но и его направление. Отбор благоприятствует "новым" фенотипам (и генотипам), а плохо приспособленные особи элими-нируются. В первую очередь при этом устраняются .особи, обладающие неадаптивными крайними фенотипами.
Одним из результатов возросшего давления отбора может быть специализация организмов к определенному образу жизни или более узкому диапазону условий среды. Это может оказаться неблаго-приятным для дальнейшей судьбы вида. Увеличение единообразия вида и его зависимости от каких-то определенных условий повышает вероятность его вымирания в случае изменения этих условий. В палеонтологической летописи встречается немало вымерших организмов, которые были крайне своеобразны и чересчур специализированы.
25.2. Как сведения о давлении отбора и образе жизни определенного паразита могут помочь искоренить его?
Из всего сказанного можно видеть, что повышенное давление отбора — это консервативный механизм, сохраняющий фенотип, лучше всего приспособленный к данным условиям среды (оптимальный фенотип).
Ослабление интенсивности отбора обычно оказывает противоположное действие. Оно может наступить при отсутствии хищников, патогенных организмов, паразитов и конкурентов или же при улучшении условий среды. Так нередко обстоит дело при проникновении вида в новую для него среду. Как полагают, именно такая ситуация благоприятствовала созданию видового разнообразия вьюрков на Галапагосских островах.
25.4. Искусственный отбор
Человек с самых ранних времен своей цивилизации применяет искусственный отбор при разведении животных и растений. Дарвин воспользовался данными по искусственному отбору, чтобы объяснить возможный механизм изменения видов, происходящего в природных популяциях, т.е. естественного отбора. Искусственный отбор основан на изоляции природных популяций и избирательном скрещивании организмов, обладающих признаками, желательными для человека. Отбор герефордской и абердин-ангусской пород крупного рогатого скота велся на количество и качество мяса, а гернзейской и джерсейской пород — на молочность. Овцы гемпширской и суффолькской пород быстро созревают и дают хорошее мясо, но они менее выносливы и менее активны в поисках пищи, чем шевиотские или шотландские черномордые овцы. Эти примеры показывают, что в одной породе невозможно объединить все признаки, необходимые для максимального экономического эффекта в любых условиях, так что программы селекции часто разрабатывают таким образом, чтобы улучшить количественно и качественно главный ценный признак данной породы.
При искусственном отборе человек создает направленное селективное давление, которое ведет к изменениям частот аллелей или генотипов в популяции. Это эволюционный механизм, приводящий к созданию новых пород, линий, сортов, рас и подвидов. Генофонды всех этих групп изолированы, но они сохраняют основную генную и хромосомную структуру, характерную для вида, к которому они все еще принадлежат.
Существует две формы искусственного отбора — инбридинг и аутбридинг.
25.4.1. Инбридинг (родственное разведение)
Инбридинг состоит в избирательном скрещивании между близко родственными особями (например, между потомками от одних и тех же родительских особей) с целью сохранения и распространения особенно желательных признаков. Инбридинг очень часто применяют при разведении таких животных, как кошки и собаки. Прежде животноводы использовали инбридинг для получения крупного рогатого скота, свиней, домашней птицы и овец, дающих много молока, мяса, яиц и шерсти соответственно, но по причинам, изложенным ниже, инбридинг не находит теперь широкого применения.
Длительный инбридинг может привести к снижению плодовитости, что создает особенно серьезные проблемы в животноводстве. Интенсивный инбридинг уменьшает изменчивость генома (общее число всех аллелей данного вида), повышая число гомозиготных генотипов за счет гетерозиготных. Для того чтобы преодолеть эти проблемы, животноводы после нескольких поколений инбридинга прибегают к аутбридингу. Например, владелец молочной фермы может проводить случку своего быка с коровами из последовательных поколений собственного стада для получения высокоудойных коров. Однако еще до того, как фермер обнаружит у своего скота признаки ослабления устойчивости к заболеваниям и снижения плодовитости, он приводит для случки другого быка или искусственно осеменяет коров спермой, приобретенной в животноводческом центре. Таким образом он вводит в стадо новые аллели и повышает гетерозиготность скрещивающейся популяции.
25.4.2. Аутбридинг (неродственное разведение)
Аутбридинг особенно полезен в растениеводстве, однако его все шире используют также в производстве мяса, яиц и шерсти. Этот метод состоит в скрещивании особей из генетически различных популяций. Аутбридинг обычно производят между представителями различных сортов или линий, а у некоторых растений — между близко родственными видами. Потомков от таких скрещиваний называют гибридами, и они превосходят по ряду признаков обе родительские формы — явление, называемое гибридной мощностью или гетерозисом. Гибридов, получа-емых при скрещивании гомозиготных родительских линий из различных популяций, называют гибридами F1; они превосходят родительские формы по, величине и количеству плодов, устойчивости к заболеваниям и скорости созревания. В случае кукурузы гибридизация привела к повышению урожаев зерна у гибридов F1 до 250% по сравнению с родительскими линиями (рис. 25.5). При двойной гибридизации гибридов, полученных от скрещивания двух инбредных линий, скрещивают между собой. Качество початков и урожайность таких гибридов с избытком окупает расходы, связанные с двухгодичной программой скрещиваний (рис. 25.6).

Рис. 25.5. Пример гибридной мощности. А. Две родительские линии кукурузы (слева и справа) и гибридная кукуруза (в середине), полученная в результате скрещивания между ними

Рис. 25.5. Пример гибридной мощности. Б. Початки этих родительских линий (вверху) и гибрида (внизу). (Фото D.F. Janes, Connecticut Agricultural Experiment Station.)

Рис. 25.6. фенотипы, образующиеся при двойной гибридизации кукурузы. Растения на правой стороне — результат скрещивания между гибридами инбредных линий (слева)
Увеличение мощности растений обусловлено повышением гетерозиготности в результате смешения генов. Например, если каждая гомозиготная родительская особь обладает некоторыми, но не всеми доминантными аллелями, определяющими мощный рост, то полученная при скрещивании таких особей гетерозигота может нести все доминантные аллели (рис. 25.7). У некоторых сортов повышение мощности может быть обусловлено не просто увеличением числа доминантных аллелей, но и каким-то взаимодействием между определенными сочетаниями аллелей в гетерозиготе. При продолжительном инбридинге гибридов F1 их мощность понижается по мере возрастания доли гомозигот (рис. 25.8).

Рис. 25.7. Простое генетическое объяснение увеличения мощности у гибридов F1

Рис. 25.8. Растения кукурузы из восьми последовательных поколений. 1 — исходный гибрид. У растений 2-8 видна постепенная утрата гибридной мощности в результате инбридинга. В трех последних поколениях, достигших гомозиготности, утрата мощности замедлилась. (Фото D. F. Jones, Connecticut Agricultural Experiment Station.)
Иногда гибридизация может сопровождаться изменением числа хромосом — полиплоидизацией, которая может приводить к возникновению новых видов (один пример этого описан в разд. 23.9.2).
25.4.3. Искусственный отбор у человека
Достигнутые за последнее время успехи в изучении структуры гена, генетического кода, механизмов наследственности и дородовой диагностики генетических дефектов создали возможности для закрепления или элиминации некоторых признаков у человека. Евгеника давно уже занимается вопросом, как можно было бы "улучшить" род человеческий путем избирательного скрещивания определенных индивидуумов. Это весьма волнующая тема, и в связи с ней возникают самые разнообразные возражения. Олдос Хаксли в своем романе "Дивный новый мир", опубликованном в 1932 г., описал то воображаемое время, когда евгеника достигнет вершины своих возможностей и будет создавать индивидуумов, соответствующих потребностям общества. Подобные идеи противоречат морали любого общества, ставящего на первое место свободу и права личности; однако можно привести немало аргументов в пользу ограниченного применения в этой области некоторых достижений генетики. В медицине получают все большее признание генетические консультации, где супругам, у которых в роду имеются генетические аномалии, разъясняют, с каким риском связано для них рождение детей. С помощью уравнения Харди-Вейнберга можно вычислить частоту носителей таких нарушений метаболизма, как фенилкетонурия, или таких болезней крови, как талассемия, серповидноклеточная анемия или гемофилия. Носителям генов того или другого из этих заболеваний можно разъяснить, какова вероятность того, что они вступят в брак с другим носителем тех же генов, и каковы шансы на то, что их дети в этом случае окажутся больными. В таких формах профилактическая медицина дает советы, а не диктует требования. Любые успехи науки, позволяющие уменьшить страдания, следует приветствовать. Такой подход становится опасным лишь в том случае, если им злоупотребляют.
25.5. Естественный отбор
Естественный отбор, постулированный Дарвином и Уоллесом, представлял собой гипотезу, основанную на исторических данных. Для Дарвина промежуток времени, необходимый для эволюционного изменения популяции, был слишком велик, чтобы такое изменение можно было наблюдать непосредственно. Происходящие в последние десятилетия изменения, связанные с промышленной, технической и медицинской революцией, создают столь сильные факторы направленного и дизруптивного отбора, что мы можем теперь наблюдать резкие изменения в генотипе и фенотипе популяций, происходящие достаточно быстро. Открытие в сороковых годах антибиотиков создало сильное давление отбора в пользу бактериальных штаммов, обладающих генетической устойчивостью к антибиотикам. Бактерии очень быстро размножаются и дают ежедневно много поколений и миллионы индивидуумов. В результате случайной мутации может появиться устойчивая клетка, потомки которой будут процветать благодаря отсутствию конкуренции со стороны других бактерий, уничтожаемых данным антибиотиком. В ответ на это приходится создавать новые антибиотики для уничтожения устойчивых бактерий, и цикл продолжается. Селективное давление создается также при использовании таких веществ, как ДДТ, для борьбы с платяной вошью и комарами или антикоагулянта уорферина для уничтожения крыс. После возникновения устойчивости она быстро распространяется во всей популяции.
Классическим примером эволюционного изменения служит реакция некоторых бабочек на направленное селективное давление, создаваемое загрязнением атмосферы в результате промышленной революции. За последние 100 лет у более чем 80 видов бабочек появились темные формы, встречающиеся теперь с различной частотой по всей Великобритании. Это явление известно под названием индустриального меланизма. До 1848 г. — все описанные формы березовой пяденицы (Biston betularia) имели бледно-кремовую окраску с черными точками и отдельными темными пятнышками (рис. 25.9,А). В 1848 г. в Манчестере была обнаружена черная форма этой бабочки, а к 1895 г. популяция березовой пяденицы в Манчестере состояла на 98% из черных бабочек. Эта черная "меланистическая" форма появилась в результате повторных случайных мутаций, причем фенотип мутантных особей обладал большим селективным преимуществом в промышленных районах по причинам, выдвинутым и проверенным д-ром Кеттлуэллом.

Рис. 25.9. Полиморфизм у березовой пяденицы Biston betularia. А. Нормальная форма В. betularia typica. Б. Меланистическая форма В. betularia carbonaria. (Е.В. Ford, Evolution studied by observation and experiment, Oxford Biology Readers, 1973, 55. Oxford Univ. Press.)
Березовые пяденицы активны в ночное время, а днем они отдыхают на стволах деревьев. Нормальная форма имеет очень эффективную покровительственную окраску, которая сливается с лишайниками, покрывающими стволы. С развитием промышленной революции двуокись серы, образующаяся при сжигании угля, вызвала в промышленных районах гибель лишайников, и в результате обнажилась темная кора деревьев, которая сделалась еще темнее из-за покрывающей ее сажи (рис. 25.10).

Рис. 25.10. Меланистическая и нормальная формы Biston betularia на стволах деревьев вблизи Бирмингема (А) и в Дорсете (Б). (С любезного разрешения д-ра Н. В. D. Kettlewell, Dep. of Zoology Univ. of Oxford.)
В пятидесятых годах нашего века Кэттлуэлл пометил определенное число светлых и темных бабочек и выпустдл их в двух местах: в загрязненном районе вблизи Бирмингема, где популяция на 90% состояла из темной формы, и в незагрязненном районе в Дорсете, где темная форма встречалась редко. С помощью световой ловушки он провел отлов меченых бабочек и получил следующие результаты:

С помощью киносъемки Кэттлуэлл показал, что малиновки и дрозды питаются этими бабочками, осуществляя форму естественного отбора, называемую избирательным выеданием, которая в данном случае оказывает селективное давление на распространение меланистической и светлой форм.
Как показывают полученные результаты, меланистическая форма, Biston betularia carbonaria, в промышленных районах обладает селективным преимуществом перед светлой формой, Biston betularia typica, тогда как светлая форма обладает преимуществом в незагрязненных районах.
Позднее было установлено, что окраска темной формы определяется доминантным меланистическим аллелем. На рис. 25.11 показано распространение этих двух форм на Британских островах.

Рис. 25.11. Распространение меланистической и нормальной форм Biston betularia на Британских островах в 1958 г. (Н. В. D. Kettlewell, Heredity, 1978, 12, 51-72.)
Распространение темных бабочек в непромышленных районах восточной части Англии объясняется преобладанием западных ветров. После того как в 1956 г. был введен в действие Акт о чистоте воздушного бассейна, доля светлых форм слегка возросла, так как в промышленных районах элиминация их отбором несколько снизилась.
25.5.1. Полиморфизм
Полиморфизм играет значительную роль в процессе естественного отбора. На примере полиморфных групп можно продемонстрировать ряд описанных в этой главе принципов, касающихся соотношения частот генотипов в популяции и изменений давления отбора. Полиморфизм можно определить как существование в пределах одной популяции двух или нескольких форм данного вида, различающихся по биохимическим, морфологическим или поведенческим признакам. Различают две формы полиморфизма: переходный и сбалансированный, или стабильный, полиморфизм.
Классическое количественное изучение сбалансированного полиморфизма провели Кейн, Карри и Шеперд на обыкновенной наземной улитке Сераеа nemoralis. У этих улиток раковина бывает желтая (если внутри находится живая улитка, раковина выглядит зеленой), различных оттенков коричневого (в том числе светло-палевая), розовая, оранжевая или красная. Устье раковины может быть темно-коричневым, розовым или белым, а на самой раковине может быть до пяти темных полос, идущих по ее контурам (рис. 25.12). Как цвет раковины, так и характер ее полос детерминированы генетически. Окраска определяется множественными аллелями, причем коричневая окраска доминирует над розовой, а обе они доминируют над желтой. Полосатость — рецессивный признак.

Рис. 25.12. Изменчивость окраски и рисунка полос у раковины Cepaea nemoralis. А — вид сверху, Б — вид сбоку. Показан постепенный переход от желтой раковины без полос (верхний ряд) до темно-коричневой полосатой (нижний ряд справа). (По Tribe, Tallan, Erant, Basic Biology Course, Book 12, Cambridge Univ. Press. 1978.)
Улиток поедают дрозды, которые переносят их на близлежащий камень и, используя его в качестве "наковальни", разбивают на нем раковину, чтобы добраться до улитки. Изучая соотношение раковин разных типов вокруг "наковален" и в самих местообитаниях улиток, Кейн, Карри и Шеперд показали, что в популяции действуют селективные факторы. В тех местах, где фон был довольно однородным, например на траве и на лесной подстилке, селективным преимуществом обладали желтые и коричневые раковины без полос, так как вблизи наковален таких раковин было меньше (рис. 25.13). Более темные полосатые раковины имели селективное преимущество там, где фон был пестрым, как, например, на пастбищах с грубыми травами и в живых изгородях. На любых участках дрозды больше всего истребляют те формы, которые хорошо заметны. Обширная популяция полиморфных улиток может занимать несколько участков, различающихся по фону. Кроме того, характер и окраска фона могут изменяться со сменой времен года. Хотя выедание хорошо заметных форм происходит непрерывно, ни одна форма не обладает абсолютным селективным преимуществом; поэтому доля каждой формы в популяции из года в год остается довольно постоянной.

Рис. 25.13. Раковины Cepaea nemoralis без полос на фоне листовой подстилки. Раковина, лежащая справа, — желтая, вверху розовая, а две раковины слева — коричневые. (По Ford, Evolution studied by observation and experiment, Oxford Biology Reader, 1973, 55, Oxford Univ. Press, 1973.)
Баланс численностей разных форм не обязательно определяется только окраской и характером полосатости. Судя по некоторым данным, сохранению равновесия при полиморфизме способствуют также физиологические факторы. В некоторых областях с сухой известковой почвой и светлой поверхностью не всегда доминируют формы с наименее заметными окраской и характером полос. Полагают, что полиморфизм у Сераеа определяется генетическим сцеплением особого типа. Гены окраски и полосатости сцеплены и образуют суперген, который действует и наследуется как одна генетическая единица. Входящие в него гены определяют признаки, обладающие такими селективными преимуществами, благодаря которым они сохраняются в популяции. Именно разнообразие аллельных форм этих генов, охраняемое благодаря широко распространенной гетерозиготности, составляет основу полиморфизма. К этому добавляется сцепление генов, детерминирующих некоторые физиологические функции, что, как полагают, тоже способствует поддержанию сбалансированного полиморфизма. Наличие в одной и той же популяции нескольких обособленных форм, доля которых слишком велика, чтобы их можно было отнести за счет повторных мутаций, называют генетическим полиморфизмом; примером служит Сераеа.
Переходный полиморфизм
Переходный полиморфизм возникает в тех случаях, когда различные формы, или морфы, существуют в популяциях, испытывающих сильное давление отбора. Частота фенотипического проявления каждой формы определяется интенсивностью отбора, как в случае меланистической и светлой форм березовой пяденицы. Переходный полиморфизм обычно наблюдается при постепенном замещении одной формы другой.
Сбалансированный полиморфизм
Сбалансированный полиморфизм создается при сосуществовании в одной популяции различных форм в стабильных условиях среды. Наиболее ярким примером служит наличие двух полов у животных и растений. Частоты генотипов разных форм сбалансированы, так как обе формы обладают равноценными селективными преимуществами. Примером сбалансированного полиморфизма служат группы крови А, В, АВ и О у человека. Частоты разных генотипов в разных популяциях могут варьировать, однако в данной популяции они остаются постоянными из поколения в поколение. Это объясняется тем, что ни один генотип не обладает селективным преимуществом перед другими. Как показывают статистические данные, у мужчин белой расы с группой крови О ожидаемая продолжительность жизни выше, чем у мужчин с другими группами крови; однако у обладателей группы О чаще, чем у других, развивается язва двенадцатиперстной кишки, которая в случае прободения может привести к смерти. Другие примеры полиморфизма — нормальное зрение и цветовая слепота у человека, касты рабочих, трутней и маток у общественных насекомых и длинно- и короткостолбчатые формы у первоцвета.
25.6. Концепция вида
Вид представляет собой самую низкую таксономическую категорию, которой еще можно дать сколько-нибудь точное определение. Существует несколько определений вида; некоторые из них приведены в табл. 25.2.

Таблица 25.2. Некоторые определения вида
Особи, принадлежащие к одному виду, редко существуют в природе в форме одной обширной популяции. Обычно вид представлен рядом небольших скрещивающихся популяций, называемых демами; каждый дем имеет собственный генофонд. Такие популяции могут занимать примыкающие друг к другу или сильно разобщенные географические области. Пространственная разобщенность популяций означает, что данный вид может сталкиваться с разнообразными условиями среды и разной интенсивностью отбора. Мутационный процесс и отбор в изолированных популяциях могут создавать различную степень фенотипической изменчивости в пределах вида, как это описано ниже.
25.6.1. Географические расы
Популяции, занимающие обширный географический ареал или в течение длительного времени разбросанные по сильно разобщенным местообитаниям, могут обладать значительными стенотипическими различиями. В основе этих различий обычно лежат приспособления к климатическим факторам. Например, непарный шелкопряд (Lymantria dispar) распространен во всей Японии и восточной части Азии. На этом пространстве существуют самые разнообразные климатические условия — от субарктических до субтропических. Различают десять географических рас шелкопряда, различающихся по срокам вылупления из яиц. Северные расы вылупляются позднее, чем южные. Как полагают, фенотипические особенности этих рас — результат влияния климатических факторов на частоты генов в их генофондах. О генетической основе такой изменчивости свидетельствует тот факт, что при помещении представителей этих рас в одинаковые условия они продолжают вылупляться в разные сроки.
25.6.2. Экологические расы (экотипы)
Популяции, приспособленные к экологически несходным местообитаниям, могут занимать смежные географические области; например, в прибрежных областях Калифорнии растение Gilia achilleaefolia представлено двумя расами. Одна раса — "солнечная" — растет на "открытых травянистых южных склонах, тогда как "теневая" раса встречается в тенистых дубовых лесах и рощах секвойи. Эти расы различаются по величине лепестков — признаку, детерминированному генетически.
25.6.3. Клины
Клиной называют постепенное изменение фенотипических признаков данного вида не протяжении его ареала. У одного вида может быть несколько клин, и они могут иметь разное направление (рис. 25.14).
Виды с выраженной фенотипической изменчивостью, связанной с частичной географической изоляцией, называют политипическими видами. Классическим примером такого вида служат чайки рода Larus (см. разд. 25.8.4).
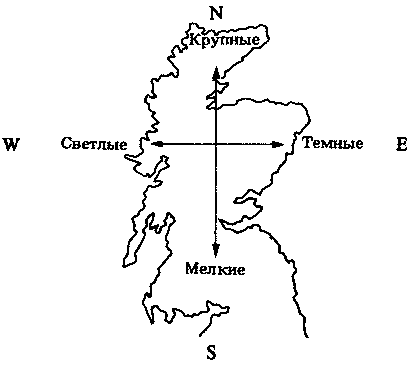
Рис. 25.14. Фенотипическая изменчивость размеров тела и окраски шерсти у лесных мышей (Apodemus) в Шотландии
Все описанные выше случаи фенотипической изменчивости отражают различную степень генетической несхожести, которая может оказывать влияние на репродуктивный потенциал членов соответствующих популяций при их встрече.
25.7. Видообразование
Видообразованием называют процесс возникновения одного или нескольких новых видов из видов, существовавших ранее. Новые виды могут возникнуть из одного вида (внутривидовое видообразование) или, как это нередко происходит у многих цветковых растений, два вида дают начало новому виду (межвидовая гибридизация). Если внутривидовое видообразование происходит в условиях пространственной разобщенности популяций, то его называют аллопатрическим видообразованием. Если же во время этого процесса популяции занимают одну и ту же географическую область, его называют симпатрическим видообразованием.
25.8. Внутривидовое видообразование
Во внутривидовом видообразовании участвует несколько факторов, однако во всех случаях непременным условием является прекращение обмена генами между популяциями; в результате каждая популяция становится генетически изолированной. Изменение частоты аллелей и генотипов в отдельных популяциях, обусловленное действием естест-венного отбора на диапазон фенотипов, создававшихся в результате мутаций и половой рекомбинации, ведет к образованию рас и подвидов. Если генетическая изоляция сохраняется в течение длительного времени, а затем подвиды вновь встречаются в одной и той же области, то они либо снова скрещиваются, либо это оказывается невозможным. В случае успешного скрещивания их все еще можно считать принадлежащими к одному виду. Невозможность скрещивания означает, что произошло видообразование и прежние подвиды следует теперь рассматривать как самостоятельные виды. Полагают, что именно таким образом могут происходить эволюционные изменения.
Начальным фактором в процессе видообразования может служить снижение интенсивности отбора в популяции. Это может привести к повышению внутривидовой изменчивости. Если новые фенотипы обладают адаптациями к условиям среды, имеющимся в краевых участках ареала, то популяция получает возможность расширить область своего распространения. В тех случаях, когда обмен генами внутри популяции не ослабевает, данный вид, проявляя локализованную фенотипическую изменчивость (экотипы), все еще сохраняет общий генофонд и продолжает оставаться единым видом. В таких ситуациях обычно создаются клины.
Видообразование может происходить только в результате возникновения преград, ведущих к репродуктивной изоляции между членами данной популяции. Репродуктивная изоляция создается изолирующими механизмами (термин, введенный генетиком Т. Добржанским) того или иного типа.
25.8.1. Изолирующие механизмы
Изолирующий механизм — это фактор, создающий и поддерживающий репродуктивную изоляцию. Репродуктивная изоляция может быть обусловлена механизмами, действующими до оплодотворения или после него. В табл. 25.3 приведена в несколько модифицированном виде классификация изолирующих механизмов, предложенная Добржанским.

Таблица 25.3. Изолирующие механизмы (по Добржанскому)
25.8.2. Аллопатрическое видообразование
Для аллопатрического видообразования характерно то, что на какой-либо его стадии существенную роль играет пространственное разобщение. Такое разобщение, создаваемое географическими барьерами (например, горными хребтами, морями или реками) или же различиями в предпочитаемых местообитаниях, может препятствовать потоку генов; организмы или их гаметы теряют возможность встретиться, и это ведет к репродуктивной изоляции. Адаптация к новым условиям и случайный дрейф генов в небольших популяциях приводят к изменениям частоты аллелей и генотипов. В результате длительного разобщения популяций между ними может возникнуть генетическая изоляция, сохраняющаяся даже в том случае, если они вновь окажутся вместе. Таким образом могут возникать новые виды. Например, разнообразие видов вьюрков семейства Goespizidae и их распространение на островах Галапагосского архипелага считают результатом аллопатрического видообразования. Как полагает Дэвид Лэк, первоначально вьюрки прибыли на острова с южноамериканского материка и здесь в отсутствие конкуренции со стороны местных видов (слабое давление отбора) произошла адаптивная радиация, приведшая к возникновению разнообразных видов, приспособленных к различным экологическим нишам; эволюция разных видов, происходившая в условиях географической изоляции, зашла так далеко, что когда впоследствии они снова встречались на некоторых островах, то могли уже сосуществовать как самостоятельные виды.
25.8.3. Симпатрическое видообразование
Наследственные различия могут накапливаться и в таких аллопатрических популяциях, которые были географически изолированы на протяжении гораздо более коротких периодов времени. Если эти популяции затем встречаются, то в зонах перекрывания могут образоваться гибриды. Например, на Британских островах обитают черная ворона (Corvus corone) и серая ворона (Corvus cor one cornix). У черной окраска целиком черная, эта форма распространена в Англии и на юге Шотландии. Серая ворона — черная с серой спинкой и брюшком и обитает в северной части Шотландии. Гибриды между черной и серой воронами занимают узкую полосу в центральных районах Шотландии (рис. 25.15). У этих гибридов низкая фертильность, и это эффективно препятствует обмену генами между популяциями черной и серой ворон.

Рис. 25.15. Зона гибридизации как барьер, препятствующий обмену генами между двумя популяциями. Благодаря наличию этой зоны, тянущейся поперек Шотландии, два вида ворон сохраняют обособленность. Между соседними популяциями часто существуют такие 'гибридные' преграды, действующие следующим образом: в тех местах, где ареалы видов А и В перекрываются, в результате межвидовых скрещиваний появляются гибриды с пониженной фертильностью; в дальнейшем вид А свободно скрещивается с гибридом АВ, а АВ с видом В, но существование гибрида АВ препятствует свободному скрещиванию между популяциями А и В
Со временем может возникнуть отбор, направленный против скрещивания между обеими формами, что приведет к образованию двух отдельных видов. Поскольку такое видообразование происходит в конечном счете в одной географической области, его называют симпатрическим видообразованием.
Симпатрическое видообразование не связано с территориальным разобщением популяций в период создания генетической изоляции. Для него необходимо развитие какого-либо механизма репродуктивной изоляции, возникающего в результате отбора в пределах географически изолированной области. Этот механизм может быть структурным, физиологическим, поведенческим или генетическим.
Симпатрическое видообразование чаще рассматривают как механизм, позволяющий объяснить, каким образом близкие виды, которые, вероятно, произошли от общего предка в результате временной изоляции, могут сосуществовать как обособленные виды в одной и той же географической области. Например, на Галапагосских островах вьюрок Саmarhyncus pauper обитает только на острове Чарлз, где он сосуществует с родственной формой С. psittacula, широко распространенной на всех центральных островах (рис. 25.16). Виды вьюрков привыборе брачного партнера руководствуются, по-видимому, размерами клюва. Диапазоны размеров клюва у С. pauper на острове Чарлз и С. psittacula на острове Альбемарль примерно одинаковы, но на острове Чарлз у С. psittacula клюв длиннее. Такое отличие уже достаточно велико для того, чтобы эти два вида, которые различаются по своему питанию, не привлекали друг друга во время брачного периода. Таким образом, виды сохраняют обособленность и способны к сосуществованию.

Рис. 25.16. Распространение двух видов вьюрков на Галапагосских островах как пример сосуществования видов после аллопатрического видообразования
25.8.4. Кольцевые клины
Симпатрическое видообразование особого типа наблюдается в местах, где две популяции, занимающие краевые участки клины, встречаются и обитают совместно в одной области, замыкая таким образом кольцо. Например, чайки рода Larus образуют непрерывную популяцию, окружающую Северный полюс кольцом между 50 и 80 ° с. ш. В это кольцо входит 10 рас или подвидов, различающихся главным образом по общим размерам тела и окраске ног, спинки и крыльев. Между всеми этими расами происходит свободный обмен генами, если не считать того места, где на Британских островах концы кольца смыкаются. Здесь, в крайних точках ареала, чайки ведут себя как два отдельных вида — серебристая чайка (Larus argentatus) и клуша (L. fuscus). Они различаются по внешнему виду, по крику и по характеру миграций и скрещиваются между собой лишь в редких случаях. Отбор, направленный против их гибридизации, происходит в условиях симпатрии.
У видов с половым размножением симпатричеcкое видообразование без географической изоляции маловероятно. Однако у организмов, размножающихся бесполым путем, в том числе у высших растений с вегетативным размножением, один мутант, достаточно отличающийся от родительской популяции, чтобы быть генетически изолированным, может симпатрически дать начало новому виду. Примером служит полиплоидия у Spartina (см. разд. 23.9.2).
25.3. Десять подвидов популяции Larus argentatus-fuscus образуют непрерывное кольцо, которое тянется от Британских островов через Скандинавию, СССР, Берингов пролив, Аляску и Канаду и возвращается к Британским островам с другой стороны. Если бы подвиды, обитающие в районе Берингова пролива и Аляски, исчезли, то какие это имело бы последствия для популяции?
25.9. Межвидовая гибридизация
Это одна из форм симпатрического видообразования, при которой новый вид возникает в результате скрещивания между особями двух разных видов. Плодовитые гибриды обычно получаются при этом только в тех случаях, когда межвидовая гибридизация приводит к хромосомной мутации, в результате которой образуется аллополиплоид (см. разд. 23.9.2). Примером служит гибрид между капустой и редькой, полученный Г. Д. Карпеченко. Происходящие при такой гибридизации генетические изменения показаны на рис. 25.17.

Рис. 25.17. Этапы создания межвидового гибрида капусты с редькой
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги