Глава 20. Размножение
Способность размножаться, т. е. производить новое поколение особей того же вида, — одна из основных особенностей живых организмов. В процессе размножения происходит передача генетического материала от родительского поколения следующему поколению, что обеспечивает воспроизведение признаков не только данного вида, но конкретных родительских особей. Для вида смысл размножения состоит в замещении тех его представителей, которые гибнут, что обеспечивает непрерывность существования вида; кроме того, при подходящих условиях размножение позволяет увеличить общую численность вида.
Каждая новая особь, прежде чем достигнуть стадии, на которой она будет способна к размножению (см. гл. 21), должна пройти ряд стадий роста и развития. Некоторые особи погибают, не достигнув репродуктивной стадии (или половозрелости) в результате уничтожения хищниками, болезней и разного рода случайных событий; поэтому вид может сохраниться лишь при условии, что каждое поколение будет производить больше потомков, чем было родительских особей, принимавших участие в размножении. Численность популяций колеблется в зависимости от баланса между размножением и вымиранием особей (разд. 12.6). Существует ряд различных стратегий размножения, каждая из которых имеет определенные преимущества и недостатки; все они будут описаны в этой главе.
20.1. Бесполое и половое размножение
Существуют два основных типа размножения — бесполое и половое. Бесполое размножение происходит без образования гамет, и в нем участвует лишь один организм. При бесполом размножении обычно образуются идентичные потомки, а единственным источником генетической изменчивости служат случайные мутации.
Генетическая изменчивость выгодна виду, так как она поставляет "сырье" для естественного отбора, а значит, и для эволюции. Потомки, оказавшиеся наиболее приспособленными к среде, будут обладать преимуществом в конкуренции с другими представителями того же вида и будут иметь больше шансов выжить и передать свои гены следующему поколению. Благодаря этому виды способны изменяться, т.е. возможен процесс видообразования (разд. 25.7). Повышение изменчивости может быть достигнуто путем смешения генов двух разных особей — процесса, называемого генетической рекомбинацией и составляющего важную особенность полового размножения; в примитивной форме генетическая рекомбинация встречается уже у некоторых бактерий (разд. 2.2.4).
20.1.1. Бесполое размножение
При бесполом размножении потомки происходят от одного организма, без слияния гамет. Мейоз в процессе бесполого размножения не участвует (если не говорить о растительных организмах с чередованием поколений; см. разд. 3.3.1), и потомки идентичны родительской особи. Идентичное потомство, происходящее от одной родительской особи, называют клоном. Члены одного клона могут быть генетически различными только в случае возникновения случайной мутации. Высшие животные не способны к бесполому размножению, однако в последнее время было сделано несколько успешных попыток клонировать некоторые виды искусственным образом; мы их рассмотрим в дальнейшем.
Существует несколько типов бесполого размножения. Подробное описание этого процесса у некоторых организмов, упоминаемых ниже, можно найти в соответствующих разделах глав 2, 3 и 4.
Деление
Делением размножаются одноклеточные организмы: каждая особь делится на две или большее число дочерних клеток, идентичных родительской клетке. Делению клетки предшествует репликация ДНК, а у эукариот — также деление ядра. В большинстве случаев происходит бинарное деление, при котором образуются две идентичные дочерние клетки. Так делятся бактерии, многие простейшие, такие как амеба или парамеция, и некоторые одноклеточные водоросли, например эвглена. При подходящих условиях это приводит к быстрому росту популяции, как было описано в разд. 2.2.4, где речь шла о бактериях.
Множественное деление, при котором вслед зарядом повторных делений клеточного ядра происходит деление самой клетки на множество дочерних клеток, наблюдается у споровиков — группы простейших, к которой относится, в частности, возбудитель малярии Plasmodium. Стадия, на которой происходит множественное деление, называется шизонтом, а сам этот процесс — шизогонией. У Plasmodium шизогония непосредственно следует за заражением хозяина, когда паразит проникает в печень. При этом получается сразу около тысячи дочерних клеток, каждая из которых способна инвазировать эритроцит и произвести путем шизогонии еще 24 дочерние клетки. Такая высокая плодовитость компенсирует большие потери из-за трудностей успешной передачи паразита от одного хозяина другому, а именно от человека организму-переносчику, т.е. малярийному комару, и в обратном направлении.
Образование спор (споруляция)
Спора — это одноклеточная репродуктивная единица обычно микроскопических размеров, состоящая из небольшого количества цитоплазмы и ядра. Образование спор наблюдается у бактерий, простейших, у представителей всех групп зеленых растений и всех групп грибов. Споры могут быть различными по своему типу и функции и часто образуются в специальных структурах. Например, у Rhizopus и Dryopleris споры образуются в спорангиях; микро-споры (пальцевые зерна) и мегаспоры (зародышевые мешки) семенных растений образуются в особых спорангиях, называемых соответственно пыльцевым мешком и семязачатком.
Нередко споры образуются в больших количествах и имеют ничтожный вес, что облегчает их распространение ветром, а также животными, главным образом насекомыми. Вследствие малых размеров спора обычно содержит лишь минимальные запасы питательных веществ; из-за того что многие споры не попадают в подходящее место для прорастания, потери спор очень велики. Главное достоинство таких спор — возможность быстрого размножения и расселения видов, в особенности грибов.
Споры бактерий служат, строго говоря, не для размножения, а для того, чтобы выжить при неблагоприятных условиях, поскольку каждая бактерия образует только одну спору. Бактериальные споры относятся к числу наиболее устойчивых: так, например, они нередко выдерживают обработку сильными дезинфицирующими веществами и кипячение в воде.
Хотя этот раздел посвящен бесполому размножению, следует указать, что существуют и половые споры — они участвуют в половом размножении. Таковы, например, зооспоры хламидомонады, иногда выполняющие функции гамет, и зигоспоры Phizopus и спирогиры. Зигоспоры — довольно крупные споры, содержащие запасы питательных веществ и окруженные защитной оболочкой; они образуются в результате полового размножения, а в периоды неблагоприятных условий могут переходить в состояние покоя.
Необходимо отметить, что один организм может производить споры более чем одного типа; например, Rhizopus образует половые и бесполые споры, а высшие растения производят бесполым путем микро- и мегаспоры.
Почкование
Почкованием называют одну из форм бесполого размножения, при которой новая особь образуется в виде выроста (почки) на теле родительской особи, а затем отделяется от нее, превращаясь в самостоятельный организм, совершенно идентичный родительскому. Почкование встречается в разных группах организмов, особенно у кишечнополостных, например у гидры (рис. 20.1), и у одноклеточных грибов, таких как дрожжи. В последнем случае почкование отличается от деления (которое тоже наблюдается у дрожжей) тем, что две образующиеся части имеют разные размеры.

Рис. 20.1. Почкование у гидры
Необычная форма почкования описана у суккулентного растения Bryophyllum — ксерофита, часто выращиваемого в качестве декоративного комнатного растения: по краям его листьев развиваются миниатюрные растеньица, снабженные маленькими корешками (рис. 20.2); эти "почки" в конце концов отпадают и начинают существовать как самостоятельные растения.

Рис. 20.2. Бесполое размножение у Bryophyllum: по краям листьев видны молодые растеньица
Размножение фрагментами (фрагментация)
Фрагментацией называют разделение особи на две или несколько частей, каждая из которых растет и образует новую особь. Фрагментация происходит, например, у нитчатых водорослей, таких как спирогира. Нить спирогиры может разорваться на две части в любом месте.
Фрагментация наблюдается также у некоторых низших животных, которые в отличие от более высокоорганизованных форм сохраняют значительную способность к регенерации из относительно слабо дифференцированных клеток. Например, тело немертин (группа примитивных червей, главным образом морских) особенно легко разрывается на много частей, каждая из которых может дать в результате регенерации новую особь. В этом случае регенерация — процесс нормальный и регулируемый; однако у некоторых животных (например, у морских звезд) восстановление из отдельных частей происходит только после случайной фрагментации. Животные, способные к регенерации, служат объектами для экспериментального изучения этого процесса; часто при этом используют свободноживущего червя планарию. Такие эксперименты помогают понять процесс дифференцировки (см. разд. 22.8).
Вегетативное размножение
Вегетативное размножение представляет собой одну из форм бесполого размножения, при которой от растения отделяется относительно большая, обычно дифференцированная, часть и развивается в самостоятельное растение. По существу вегетативное размножение сходно с почкованием. Нередко растения образуют структуры, специально предназначенные для этой цели: луковицы, клубнелуковицы, корневища, столоны и клубни. Некоторые из этих структур служат также для запасания питательных веществ, что позволяет растению пережить периоды неблагоприятных условий, таких как холода или засуха. Запасающие органы позволяют растению переживать зиму и давать в следующем году цветки и плоды (двулетние растения) или выживать в течение ряда лет (многолетние растения). К таким органам, называемым зимующими, относятся луковицы, клубнелуковицы, корневища и клубни.
Зимующими органами могут быть также стебли, корни или целые побеги (почки), однако во всех случаях содержащиеся в них питательные вещества создаются главным образом в процессе фотосинтеза, происходящего в листьях текущего года. Образовавшиеся питательные вещества переносятся в запасающий орган, а затем обычно превращаются в какой-либо нерастворимый резервный материал, например крахмал. При наступлении неблагоприятных условий надземные части растения отмирают, а подземный зимующий орган переходит в состояние покоя. В начале следующего вегетационного периода запасы питательных веществ мобилизуются с помощью ферментов: почки пробуждаются, и в них начинаются процессы активного роста и развития за счет запасенных питательных веществ. Если прорастает более одной почки, то можно считать, что осуществилось размножение. Эта последовательность событий тесно согласована со сменой времен года, поскольку она регулируется такими внешними факторами, как длина светового дня (фотопериод) и температура, глубокое влияние которых на рост и развитие было описано в гл. 15.
Луковица. Это видоизмененный побег, имеющийся, например, у лука (Allium), нарцисса (Narcissus) и тюльпана (Tulipa). Она служит как зимующим органом, так и органом вегетативного размножения.
Луковица состоит из очень короткого стебля и мясистых листьев, содержащих запасные питательные вещества. Снаружи она покрыта бурыми пленчатыми листьями — остатками прошлогодних листьев, запасные вещества которых были израсходованы. Луковица содержит одну или несколько дочерних луковиц (детки или зубки); каждая из них может образовать побег, который к концу вегетационного периода дает новую луковицу, и если их будет несколько, то это уже вегетативное размножение. Корни у луковиц придаточные, т.е. отходят непосредственно от стебля, а главного корня нет.
Типичная луковица изображена на рис. 20.3.

Рис. 20.3. Схематический продольный разрез через покоящуюся луковицу
Клубнелуковица. Это короткий, вздутый вертикальный подземный стебель, как, например, у шафрана (Crocus) или гладиолуса (Gladiolus). Клубнелуковицы служат как запасающими органами, так и органами вегетативного размножения.
Клубнелуковица состоит из вздутого основания стебля, окруженного защитными пленчатыми листьями; в отличие от луковицы здесь нет мясистых листьев. Пленчатые листья — это остатки прошлогодних надземных листьев. Корни придаточные. К концу вегетационного периода они укорачиваются и втягивают новую клубнелуковицу в почву. Клубнелуковица (так же как луковица) содержит одну или несколько деток, которые могут обеспечить вегетативное размножение.
Типичная клубнелуковица показана на рис. 20.4.
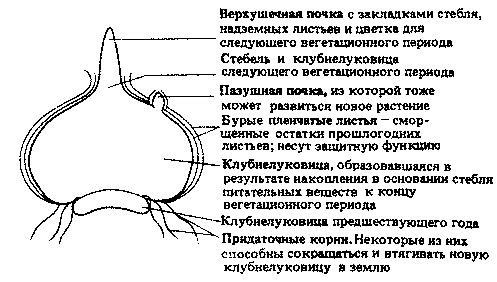
Рис. 20.4. Схематический продольный разрез через покоящуюся клубнелуковицу
Корневище. Оно представляет собой подземный стебель, растущий горизонтально. У одних растений, например у ириса и соломоновой печати (Polygonatum), корневище короткое и вздутое, содержащее запасные питательные вещества, а у других, таких как пырей ползучий (Аgropyron repens), мята (Mentha) и астры (Aster spp.), оно длинное и тонкое. Корневище обычно служит зимующим органом, а также органом вегетативного размножения.
Корневище несет листья, почки и придаточные корни. Листья могут быть либо пленчатые или чешуйчатые (мелкие, тонкие, беловатого или коричневатого цвета), как у пырея, либо только зеленые надземные, как у ириса, либо того и другого типа, как, например, у соломоновой печати, у которой листья обоих типов растут и на надземных побегах. Корневище ириса изображено на рис. 20.5.

Рис. 20.5. Схема строения корневища ириса
Столон. Это ползучий горизонтальный стебель, стелющийся на поверхности почвы, как, например, у ежевики (Rubus), крыжовника (Grossularia), черной и красной смородины (Ribes spp.). Столон не служит зимующим органом. Корни придаточные, отходящие от узлов.
Схема строения типичного столона представлена на рис. 20.6.

Рис. 20.6. Обобщенная схема строения столона
Усы (плети). Это разновидность столонов, которые быстро растут в длину, как, например, у земляники (Fragaria) или у лютика ползучего (Ranunculus repens).
Ус несет пленчатые листья с пазушными почками, которые дают начало придаточным корням и новым растениям. После укоренения новых растений прежние усы в конце концов разрушаются. В роли уса может выступать главный стебель; ус может также расти от одной из нижних пазушных почек на главном стебле, как показано на рис. 20.7. У земляники пленчатые листья и пазушные почки есть у каждого узла, но корни и надземные листья образует лишь каждый второй узел. Все пазушные почки могут дать начало новым усам.

Рис. 20.7. Строение уса земляники
Клубень. Это подземный запасающий орган, раздутый в результате накопления питательных веществ и способный перезимовывать. Клубни живут только один год, а затем, после того как их содержимое будет использовано за время вегетационного периода, они ссыхаются. К концу вегетационного периода образуются новые клубни, но они возникают не из старых клубней (в отличие от клубнелуковиц, возникающих из прежних клубнелуковиц).
Стеблевые клубни представляют собой структуры, образующиеся на концах тонких корневищ, как у картофеля (Solarium tuberosum) и топинамбура (Helianthus tuberosum). На их стеблевое происхождение указывает наличие пленчатых листьев и почек в их пазухах. В следующем вегетационном периоде каждая такая почка может дать начало новому растению.
Корневые клубни — это вздувшиеся придаточные корни, как, например, у георгины (Dahlia) и у Ranunculus ficaria. Новые растения развиваются из пазушных почек у основания старого стебля.
Стеблевой клубень картофеля изображен на рис. 20.8, а корневые клубни георгины — на рис. 20.9.

Рис. 20.8. Стеблевой клубень картофеля

Рис. 20.9. Корневые луковицы георгины
Мясистые стержневые корни. Стержневой корень — это главный корень, развивающийся из первичного корешка, т.е. первого корня зародыша. Корневая система стержневого типа характерна для двудольных растений. У некоторых растений, таких как морковь (Daucus), пастернак (Pastinaca), брюква (Brassica napus), репа (Brassica rара) и редька (Каphanus sativus), главный корень может утолщаться за счет развития паренхимной ткани, содержащей запасные питательные вещества. Вместе с почками, расположенными у основания старого стебля непосредственно над стержневым корневым, такие корни образуют зимующие органы и органы вегетативного размножения. Мясистые стержневые корни двух типов показаны на рис. 20.10.

Рис. 20.10. Стержневые корни моркови и репы
Мясистые стержневые корни характерны для двулетних растений, у которых в первый год происходит вегетативный рост. Затем они переживают зиму за счет подземного запасающего органа, а на второй год образуют цветки и семена и к концу года отмирают.
Помимо описанных выше специализированных органов вегетативного размножения новое растение могут регенерировать и некоторые другие, неспециализированные органы, если отделить их от родительского растения, например листья суккулента Sedum. Одной из форм вегетативного размножения можно считать также размножение черенками и отводками, применяемое в плодоводстве и цветоводстве.
Черенки и отводки — это части растения, которые в подходящих условиях пускают корни и образуют листья, превращаясь в самостоятельные растения. Таким образом можно искусственно размножать нужные сорта, которые при этом не изменяются. Нередко укоренение стимулируют добавлением ростового гормона. Для вегетативного размножения с успехом используют побеги пеларгонии и колеуса, веточки ивы (Salix spp.) и Forsythia, а также листья бегонии и узамбарской фиалки (Saintpaulia ionantha).
Другим важным и широко используемым способом искусственного вегетативного размножения служат прививки. Прививка состоит в пересадке одного растения (побега или почки) на нижнюю часть побега другого растения. Пересаженную часть растения — донора называют привоем, а реципиента — подвоем. Подвой обрезают над местом прививки. Получающееся в результате растение обычно обладает корневой системой подвоя и побегом (в том числе цветками и плодами) привоя. Этот метод применяется в широких масштабах для размножения розовых кустов и плодовых деревьев, в особенности яблонь. Он имеет два преимущества — позволяет сочетать в одном растении желательные признаки двух разных сортов или видов и дает возможность быстро получать в больших количествах новые комбинации привой/подвой для продажи. Иногда удается получить прививки нескольких привоев на данном подвое; примером служат яблони, у которых на одном дереве вырастают десертные плоды и плоды, предназначенные для варки варенья и консервирования.
Клонирование высших растений и животных
Как уже говорилось, получение идентичных потомков при помощи бесполого размножения называют клонированием. В начале шестидесятых годов были разработаны методы, позволяющие успешно клонировать некоторые высшие растения и животных. Эти методы возникли в результате попыток доказать, что ядра зрелых клеток, закончивших свое развитие, содержат всю информацию, необходимую для кодирования всех признаков организма, и что специализация клеток обусловлена включением и выключением определенных генов, а не утратой некоторых из них (см. разд. 22.7). Первый успех был достигнут проф. Стюардом из Корнелльского университета, который показал, что, выращивая отдельные клетки корня моркови (ее съедобной части) в среде, содержащей нужные питательные вещества и гормоны, можно индуцировать процессы клеточного деления, приводящие к образованию новых растений моркови.
Вскоре после этого Гёрдон, работавший в Оксфордском университете, впервые сумел добиться клонирования позвоночного животного. Позвоночные в естественных условиях клонов не образуют; однако, пересаживая ядро, взятое из клетки кишечника лягушки, в яйцеклетку, собственное ядро которой предварительно было разрушено путем облучения ультрафиолетом, Гёрдону удалось вырастить головастика, а затем и лягушку, идентичную той особи, от которой было взято ядро (рис. 20.11).

Рис. 20.11. Клон шпорцевых лягушек (Xenopus laevis), полученный путем трансплантации ядер. А. От скрещивания между двумя мутантными лягушками-альбиносами был получен зародыш (донор); на стадии хвостовой почки его клетки были диссоциированы и выделенные ядра пересажены в неоплодотворенные яйца самки дикого типа (реципиента), ядра которых были разрушены УФ-облучением

Рис. 20.11. Клон шпорцевых лягушек (Xenopus laevis), полученный путем трансплантации ядер. Б. Группа из 30 лягушек, полученных в результате 54 таких пересадок; все они — самки — альбиносы
Такого рода эксперименты не только доказывают, что дифференцированные (специализированные) клетки содержат всю информацию, необходимую для развития целого организма, но и позволяют рассчитывать, что подобные методы можно будет использовать для клонирования позвоночных, стоящих на более высоких ступенях развития, в том числе и человека. Клонирование нужных животных, например племенных быков, скаковых лошадей и т.п., может оказаться столь же выгодным, как и клонирование растений, которое, как было сказано, уже производится. Однако применение методов клонирования к человеку сопряжено с серьезными проблемами нравственного порядка. Теоретически можно создать любое число генетически тождественных копий данного мужчины или данной женщины. На первый взгляд может показаться, что таким образом можно было бы воспроизводить талантливых ученых или деятелей искусства. Однако надо помнить, что степень влияния, оказываемого на развитие средой, еще не вполне ясна, а между тем любая клонируемая клетка должна снова пройти через все стадии развития, т.е. в случае человека-стадии зародыша, плода, младенца и т.д.
20.1.2. Половое размножение
При половом размножении потомство получается в результате слияния генетического материала гаплоидных ядер. Обычно эти ядра содержатся в специализированных половых клетках-гаметах; при оплодотворении гаметы сливаются, образуя диплоидную зиготу, из которой в процессе развития получается зрелый организм. Гаметы гаплоидны — они содержат один набор хромосом, полученный в результате мейоза; они служат связующим звеном между данным поколением и следующим (при половом размножении цветковых растений сливаются не клетки, а ядра, но обычно эти ядра тоже называют гаметами.)
Мейоз — важный этап жизненных циклов, включающих половое размножение, так как он ведет к уменьшению количества генетического материала вдвое. Благодаря этому в ряду поколений, размножающихся половым путем, это количество остается постоянным, хотя при оплодотворении оно каждый раз удваивается. Во время мейоза в результате случайного расхождения хромосом (независимое распределение) и обмена генетическим материалом между гомологичными хромосомами (кроссинговер) возникают новые комбинации генов, попавших в одну гамету, и такая перетасовка повышает генетическое разнообразие (см. разд. 22.3). Слияние содержащихся в гаметах гаплоидных ядер называют оплодотворением или сингамией; оно приводит к образованию диплоидной зиготы, т. е. клетки, содержащей по одному хромосомному набору от каждого из родителей. Это объединение в зиготе двух наборов хромосом (генетическая рекомбинация) представляет собой генетическую основу внутривидовой изменчивости. Зигота растет и развивается в зрелый организм следующего поколения. Таким образом, при половом размножении в жизненном цикле происходит чередование диплоидной и гаплоидной фаз, причем у разных организмов эти фазы принимают различные формы (см. рис. 20.13).
Гаметы обычно бывают двух типов — мужские и женские, но некоторые примитивные организмы производят гаметы только одного типа (см. в разд. 20.2 об изогамии, анизогамии и оогамии). У организмов, образующих гаметы двух типов, их могут производить соответственно мужские и женские родительские особи, а может быть и так, что у одной и той же особи имеются и мужские, и женские половые органы. Виды, у которых существуют отдельные мужские и женские особи, называются раздельнополыми; таковы большинство животных и человек. Среди цветковых растений тоже есть раздельнополые виды; если у однодомных видов мужские и женские цветки образуются на одном и том же растении, как, например, у огурца (Cucumis) и лещины (Corylus), то у двудомных одни растения несут только мужские, а другие — только женские цветки, как у остролиста (Ilex) или у тиса (Taxus).
Гермофродитизм
Виды, у которых одна и та же особь способна производить и мужские, и женские гаметы, называют гермафродитными или двуполыми. К их числу относятся многие простейшие, в том числе парамеция, такие кишечнополостные, как Obelia, плоские черви, например солитер (Taenia), олигохеты, например дождевой червь (lumbricus), ракообразные, например морской желудь (Balanus), такие моллюски, как улитка (Helix), некоторые рыбы и ящерицы, а также большинство цветковых растений. Гермафродитизм считается самой примитивной формой полового размножения и свойствен многим примитивным организмам. Он представляет собой приспособление к сидячему, малоподвижному или паразитическому образу жизни. Одно из преимуществ гермафродитизма состоит в том, что он делает возможным самооплодотворение, а это весьма существенно для некоторых эндопаразитов, таких как солитер, ведущих одиночное существование. Однако у большинства гермафродитных видов в оплодотворении участвуют гаметы, происходящие от разных особей, и у них имеются многочисленные генетические, морфологические и физиологические адаптации, препятствующие самооплодотворению и благоприятствующие перекрестному оплодотворению. Например, у многих простейших самооплодотворение предотвращается генетической несовместимостью, у многих цветковых растений — строением андроцея и гинецея, а у многих животных — тем, что яйца и спермии образуются у одной и той же особи в разное время.
Партеногенез
Партеногенез — одна из модификаций полового размножения, при которой женская гамета развивается в новую особь без оплодотворения мужской гаметой. Партеногенетическое размножение встречается как в царстве животных, так и в царстве растений, и преимущество его состоит в том, что в некоторых случаях оно повышает скорость размножения.
Существует два вида партеногенеза — гаплоидный и диплоидный, в зависимости от числа хромосом в женской гамете. У многих насекомых, в том числе у муравьев, пчел и ос, в результате гаплоидного партеногенеза в пределах данного сообщества возникают различные касты организмов. У этих видов происходит мейоз и образуются гаплоидные гаметы. Некоторые яйцеклетки оплодотворяются, и из них развиваются диплоидные самки, тогда как из неоплодотворенных яйцеклеток развиваются фертильные гаплоидные самцы. Например, у медоносной пчелы (Apis mellifera) матка откладывает оплодотворенные яйца (2n = 32), которые, развиваясь, дают самок (маток или рабочих особей), и неоплодотворенные яйца (n = 16), которые дают самцов (трутней), производящих спермии путем митоза, а не мейоза. Развитие особей этих трех типов у медоносной пчелы схематически представлено на рис. 20.12. Такой механизм размножения у общественных насекомых имеет адаптивное значение, так как позволяет регулировать численность потомков каждого типа.

Рис. 20.12, Схема, иллюстрирующая роль партеногенеза в жизненном цикле семьи медоносной пчелы
У тлей происходит диплоидный партеногенез, при котором ооциты самки претерпевают особую форму мейоза без расхождения хромосом (см. разд. 22.3) — все хромосомы переходят в яйцеклетку, а полярные тельца не получают ни одной хромосомы. Яйцеклетки развиваются в материнском организме, так что молодые самки рождаются вполне сформировавшимися, а не вылупляются из яиц. Такой процесс называется живорождением. Он может про-должаться в течение нескольких поколений, особенно летом, до тех пор пока в одной из клеток не произойдет почти полное нерасхождение, в результате чего получается клетка, содержащая все пары аутосом и одну Х-хромосому. Из этой клетки партеногенетически развивается самец. Эти осенние самцы и партеногенетические самки производят в результате мейоза гаплоидные гаметы, участвующие в половом размножении. Оплодотворенные самки откладывают диплоидные яйца, которые перезимовывают, а весной из них вылупляются самки, размножающиеся партеногенетически и рождающие живых потомков. Несколько партеногенетических поколений сменяются поколением, возникающим в результате нормального полового размножения, что вносит в популяцию генетическое разнообразие в результате рекомбинации. Главное преимущество, которое дает тлям партеногенез, — это быстрый рост численности популяции, так как при этом все ее половозрелые члены способны к откладке яиц. Это особенно важно в периоды, когда условия среды благоприятны для существования большой популяции, т.е. в летние месяцы.
Партеногенез широко распространен у растений, где он принимает различные формы. Одна из них — апомиксис — представляет собой партеногенез, имитирующий половое размножение. Апомиксис наблюдается у некоторых цветковых растений, у которых диплоидная клетка семязачатка — либо клетка нуцеллуса, либо мегаспора — развивается в функциональный зародыш без участия мужской гаметы. Из остального семязачатка образуется семя, а из завязи развивается плод. В других случаях требуется присутствие пыльцевого зерна, которое стимулирует партеногенез, хотя и не прорастает; пыльцевое зерно индуцирует гормональные изменения, необходимые для развития зародыша, и на практике такие случаи трудно отличить от настоящего полового размножения.
20.1.3. Происхождение полового процесса
Любые рассуждения о событиях, происходящих на неведомом нам этапе истории жизни, носят чисто умозрительный характер, и вытекающие из них выводы следует считать лишь предположительными. Именно так обстоит дело с происхождением полового процесса. Предполагается, что бесполое размножение представляет собой примитивную форму вопроизведения как способа "копирования" генетически идентичных поколеней. На более позднем этапе возникли механизмы, делающие возможным обмен нуклеиновыми кислотами между особями. Быть может, при этом происходило слияние целых организмов, за которым следовал мейоз; и в самом деле, такой процесс мог предшествовать образованию гамет и их слиянию. Принято считать, что способы обмена генетическим материалом, наблюдаемые у бактерий (см. разд. 2.2.4), представляют собой ранний этап развития полового процесса. Преимущество, которое дает фенотипическая изменчивость, обусловленная мейозом и генетической рекомбинацией, несомненно, сыграло важную роль в развитии более сложных форм жизни и типов гамет — от идентичных гамет (изогамет) к гетерогаметной стадии с подвижными мужскими гаметами (спермиями и антерозоидами) и неподвижными женскими гаметами (яйцеклетками). Тот факт, что половое размножение мы находим во всех крупных группах организмов, позволяет думать, что оно обладает преимуществом перед бесполым, которое, будучи редким у животных, стоящих по своей организации выше плоских червей, сохраняется у растений: в жизненных циклах большинства растений наблюдается чередование бесполого и полового способов размножения (разд. 20.2). Одна из причин этого состоит в том, что у неподвижных организмов обмен гаметами затруднен и они во многих случаях предпочитают бесполое размножение. Кроме того, при бесполом размножении родительские особи, обосновавшиеся в определенном месте, облегчают потомкам решение нелегкой экологической проблемы — нахождение подходящего местообитания. Однако при этом возникают другие проблемы, создаваемые перенаселением, что способствует ограничению размеров растительных популяций.
Некоторые из наиболее существенных и типичных черт бесполого и полового размножения перечислены в табл. 20.1.

Таблица 20.1. Сравнение бесполого и полового размножения[5]
20.1.4. Разнообразие жизненных циклов
Последовательность стадий развития, через которые проходят представители данного вида от зиготы одного поколения до зиготы следующего, называют жизненным циклом. Сложность жизненных циклов у разных организмов различна; в некоторых случаях жизненный цикл включает два и больше поколений, различающихся по своей морфологии (внешнему виду) и способу размножения, — явление, называемое иногда чередованием поколений. Применение этого термина лучше ограничить наземными растениями и некоторыми эволюционно продвинутыми водорослями, у которых наблюдается чередование диплоидного спорообразующего поколения, называемого спорофитом, и гаплоидного гаметофита, образующего гаметы (см. разд. 3.3.1). Чередование бесполого и полового поколений имеет место также у некоторых кишечнополостных, но в этом случае оба поколения диплоидны, а гаплоидная стадия представлена только гаметами. Такую форму чередования поколений иногда называют метагенезом (см. разд. 4.4.3). В жизненном цикле этих кишечнополостных разные формы особей данного вида сменяют одна другую — явление, известное под названием циклического полиморфизма. Например, в жизненном цикле обелии последовательно сменяются три различные формы — две полипоидные и одна медузоидная (рис. 4.11). Полиморфизм встречается и у некоторых других организмов, например у первоцвета (Primula), образующего цветки двух типов — длинностолбчатые и короткостолбчатые.
Жизненные циклы паразитов часто бывают весьма сложными и включают несколько поколений. Каждое поколение приспособлено к определенной жизненной ситуации — либо к существованию в организме определенного хозяина, либо к переходу от одного хозяина к другому (примером служит жизненный цикл печеночной двуустки, см. разд. 4.5.3). Добавочные поколения, возникающие путем бесполого размножения (полиэмбриония), обеспечивают увеличение численности данного вида. Некоторые типичные жизненные циклы представлены на рис. 20.13.

Рис. 20.13. Схемы различных часто встречающихся жизненных циклов. Обратите внимание, что в жизненном цикле каждого типа мейоз происходит только один раз. На схемах А, Б и Г бесполое размножение полового организма не показано, хотя у ряда видов оно возможно, n-гаплоид, 2n-диплоид
A. Взрослый (половой) организм гаплоидный. Диплоидна только зигота. Первое деление ядра в прорастающей зиготе происходит путем мейоза и приводит к возврату в гаплоидное состояние. Гаметы образуются не в результате мейоза. Примеры: Chlamydomonas, Spirogyra, Rhizopus.
Б. Гаплоидны только гаметы; они образуются в результате мейоза. Примеры: Fucus, позвоночные и большинство других животных.
B. В жизненном цикле участвуют три морфологически различные формы (полиморфизм). Все они диплоидные. Диплоидное половое поколение чередуется с диплоидным бесполым поколением. Единственные гаплоидные клетки это гаметы; они образуются путем мейоза. Пример: Obelia.
Г. Чередование гаплоидных и диплоидных поколений. Гаметы образуются не в результате мейоза. Примеры: Laminaria (бурая водоросль), все наземные растения, т. е. мхи, папоротники и семенные растения.
Ввиду того что вопрос об относительной роли митоза и мейоза в бесполом и половом размножении иногда вызывает затруднения, мы еще раз рассмотрим здесь значение каждого из этих процессов в жизненном цикле. Мейоз происходит только в тех жизненных циклах, которые включают половое размножение. Зигота, образующаяся в результате слияния ядер двух гаплоидных гамет, содержит диплоидное число хромосом и начинает следующий жизненный цикл. Если на какой-либо стадии этого цикла не произойдет мейоза, то ядра образующихся гамет будут диплоидными, и новая зигота в таком случае окажется тетраплоидной (будет содержать четыре набора хромосом). Таким образом, мейоз необходим для того, чтобы предотвратить удвоение хромосом в каждом поколении в жизненных циклах, включающих половое размножение. Мейоз не обязательно происходит непосредственно перед образованием гамет (см. рис. 20.13,А и Г). Еще одно преимущество мейоза-то, что он создает изменчивость в результате независимого распределения хромосом и кроссинговера (разд. 22.3). Эти процессы непосредственно (жизненные циклы Б и В на рис. 22.3) или косвенным образом (циклы А и Г) приводят к различию между гаметами. В особом случае чередования поколений (жизненный цикл Г) мейоз происходит при образовании спор, а не гамет. Поэтому здесь создаются различия между спорами и между развивающимися из них гаметофитами. Эта изменчивость передается ядрам гамет, образующихся в гаметофитах путем митоза. Во всех случаях, однако, мейоз составляет основу полового размножения. В жизненных циклах А, Б и В, т. е. в циклах, не связанных с чередованием поколений, в результате бесполого размножения образуются генетически идентичные потомки, и это происходит без мейоза.
20.1 А. На основании всего сказанного в разд. 20.1 укажите, какое из сформулированных ниже утверждений верно.
а) Бесполое размножение всегда приводит к образованию генетически идентичных потомков.
б) Гаметы всегда гаплоидны.
в) Гаметы всегда образуются в процессе мейоза.
г) В результате мейоза всегда образуются гаплоидные, а в результате митоза — диплоидные клетки.
д) Митоз происходит только в диплоидных клетках.
Б. Приведите примеры, опровергающие четыре остальных утверждения.
20.2. Половое размножение у растений
У всех зеленых растений, обитающих на суше, а также у некоторых продвинутых в эволюционном отношении водорослей наблюдается смена поколений, при которой гаплоидное гаметофитное поколение чередуется с диплоидным спорофитным, как это показано на рис. 20.13,Г. Половое размножение происходит на стадии гаметофита, который образует гаметы путем митоза.
Все прочие зеленые растения относятся к водорослям, и у них можно встретить разнообразные типы полового размножения (см. разд. 3.2.3). При простейших типах (обычно их считают примитивными) все гаметы идентичны, а процесс размножения называют изогамией; при более сложных (или эволюционно продвинутых) типах наблюдается анизогамия-гаметы различаются по величине и поведению; при крайней форме анизогамии — оогамии — подвижная мужская гамета подплывает к неподвижной женской.
На протяжении миллионов лет после выхода из воды на сушу и у растений, и у животных вырабатывались разнообразные приспособления, необходимые для наземной жизни. Наиболее продвинувшиеся в эволюционном отношении группы растений и животных, а именно цветковые растения и млекопитающие, в значительной степени обязаны своим успехом адаптациям, касающимся полового размножения, и здесь между этими двумя группами можно усмотреть некоторые аналогии. Например, и у тех и у других родительские особи обеспечивают оплодотворение, не полагаясь на встречу гамет, выпущенных в воду (как это делают, например, мхи или амфибии), и, кроме того, обеспечивают развивающемуся зародышу защиту и питание. Одно из главных различий состоит в том, что у цветковых растений гаметы производит гаметофитное поколение путем митоза, тогда как у млекопитающих нет чередования поколений и гаметы образуются у диплоидных родительских особей в результате мейоза. Как мы увидим, однако, гаметофит у цветковых растений крайне редуцирован.
20.2.1. Жизненный цикл цветковых растений (покрытосеменных)
Адаптации цветковых растений к жизни на суше были описаны в гл. 3. Главные адаптации в области размножения — это образование семян и плодов, обеспечивающих питание и защиту зародышам и способствующих расселению растений, отсутствие свободноплавающих мужских гамет и крайняя редукция полового гаметофитного поколения. Мужские гаметы, заключенные в пыльцевых зернах, переносятся на женские органы растения (процесс, называемый опылением), а затем происходит прорастание пыльцевой трубки, содержащей мужские ядра.
Жизненный цикл цветковых растений схематически представлен на рис. 3.37, где он сопоставляется с жизненными циклами, типичными для других групп растений. В нем все еще сохраняется чередование поколений, показанное в упрощенном виде на рис. 20.13,Г. Поскольку гаметофитное поколение почти исчезло и уже не представлено свободноживущей особью, было бы трудно заметить, что чередование поколений имеет место, если бы не возможность провести сравнение с более примитивными предками. Жизненный цикл цветкового растения подробно описан ниже; строго говоря, он складывается из бесполого размножения доминирующего спорофита (цветковое растение как таковое) и полового размножения гаметофита.
20.2.2. Строение и функции цветка
Термин "цветковые растения", как обычно называют покрытосеменные, указывает на признак, присущий только этой группе, — наличие цветков. Цветки — это репродуктивные органы, эволюционное происхождение которых неясно; некоторые авторы полагают, однако, что цветок построен из высокоспециализированных листьев и аналогичен шишке голосеменных, так как несет спорообразующие структуры. Цветки можно считать органами как бесполого, так и полового размножения — бесполого потому, что они производят споры (пыльцевые зерна и зародышевые мешки), а полового — потому что в дальнейшем в спорах образуются гаметы. Обычно цветки называют просто органами полового размножения. В областях с четко выраженной сменой времен года цветение приурочено к определенному сезону. Регуляция цветения гормонами и факторами среды описана в разд. 15.5.
Расположение частей цветка на цветоложе — верхней части цветоноса (цветоножки) — бывает спиральным или мутовчатым. На рис. 20.14 представлены обобщенная схема строения цветка и различные типы соцветий, т. е. групп объединенных цветков, а ниже мы даем объяснение некоторых терминов, используемых при описании цветков.

Рис. 20.14. А Схема строения цветка (продольный разрез). Собирательные термины указаны стрелками; например, 'лепесток → ВЕНЧИК' означает, что совокупность лепестков называют венчиком. Б. Расположение цветков по одному или группами (соцветия). Показаны некоторые обычные типы соцветий, которые делятся на ботрические (или рацемозные — кистевидные) и цимозные.
Части цветка
Соцветием называют совокупность цветков, расположенных на конце одного стебля — ножке соцветия (рис. 20.14,Б). Вероятно, большое число цветков, собранных вместе, лучше привлекает насекомых — опылителей, чем маленький одиночный цветок. Дистальную часть цветоножки, на которой располагаются околоцветник, гинецей и андроцей, называют цветоложем. Цветок называют подпестичным (гипогинным), если тычинки и околоцветник расположены под гинецеем (рис. 20.14,А), надпестичным (эпигинным), если они расположены над гинецеем, и околопестичным (перигинным), если гинецей находится в центре уплощенного или вогнутого цветоложа, а тычинки и околоцветник прикреплены к его верхнему краю (см. также описание верхней и нижней завязи, данное ниже).
Околоцветник состоит из двух мутовок листовидных частей, называемых чашелистиками и лепестками. У однодольных обе мутовки обычно одинаковые, как, например, у нарцисса (Narcissus), тюльпана (Tulipa) и пролески Endymion (Scilla) non-scripta. У двудольных мутовки часто отличаются одна от другой: наружная, состоящая из чашелистиков, называется чашечкой, а внутренняя, состоящая из лепестков, — венчиком.
Чашелистики, образующие чашечку, — это обычно листовидные образования зеленого цвета, которые служат для защиты заключенным внутри цветочным бутонам. Иногда они могут быть ярко окрашены и похожи на лепестки; в таких случаях они тоже участвуют в привлечении насекомых — опылителей.
Свободнолистная чашечка состоит из свободных (несросшихся) чашелистиков, а спайнолистная — из чашелистиков, которые хотя бы частично слились в трубку.
Венчиком называют совокупность лепестков цветка. У растений, опыляемых насекомыми, лепестки обычно крупные и яркоокрашенные — они привлекают насекомых к цветкам. У ветроопыляемых растений лепестки чаще мелкие и зеленые, а иногда отсутствуют вовсе.
Свободнолепестный венчик состоит из свободных (несросшихся) лепестков, как, например, у горошка (Pisum), розы (Rosa) и лютика (Ranunculus); у спайнолепестного венчика лепестки хотя бы частично слиты в трубочку, как, например, у наперстянки (Digitalis), первоцвета (Primula), одуванчика (Taraxacum) и яснотки (Lamium album).
Андроцей — это совокупность тычинок, образующих мужские репродуктивные органы цветка. Каждая тычинка состоит из пыльника и тычиночной нити. Пыльник содержит пыльцевые мешки, в которых образуется пыльца. Тычиночная нить содержит проводящий пучок, по которому в пыльник поступают питательные вещества и вода.
Гинецей, или пестик, — это совокупность плодолистиков, или карпелей, образующих женские репродуктивные органы цветка. Плодолистик состоит из рыльца, столбика и завязи. Столбик поддерживает рыльце в таком положении, чтобы на него могла попадать пыльца во время опыления. Завязь представляет собой вздутое полое основание плодолистика, содержащее один или несколько семязачатков. Семязачатки — это структуры, в которых развиваются зародышевые мешки и которые после оплодотворения превращаются в семена. Каждый семязачаток прикреплен к стенке завязи короткой ножкой-фуникулусом, а место его прикрепления носит название плаценты.
Плодолистики цветка могут оставаться свободными, т.е. не связанными друг с другом, образуя апокарпный гинецей, как у лютика, или же слиты в единую структуру — синкарпный гинецей, как у яснотки. Завязь, образуемая слившимися плодолистиками, может иметь одну камеру, или гнездо (одно-гнездная завязь), или же несколько (многогнездная завязь); в последнем случае каждый из составляющих ее плодолистиков образует по одному гнезду. Столбики синкарпного гинецея могут сливаться или сохранять обособленность.
Верхняя завязь — завязь, расположенная на цветоложе над другими частями цветка, как в подпестичном или околопестичном цветке; нижняя завязь расположена на цветоложе под другими частями цветка, как в надпестичном цветке.
Нектарники — это железистые образования, выделяющие нектар — сахаристую жидкость, которая привлекает животных-опылителей (обычно это насекомые, но в тропических областях опылителями могут быть также птицы и летучие мыши).
Термины, перечисленные ниже, приложимы как к растению в целом, так и к цветку.
Гермафродитные (обоеполые) растения: мужские и женские половые органы образуются на одном и том же растении.
Однодомные растения: мужские и женские цветки растут по отдельности на одном и том же растении, как, например, у дуба (Quercus), лещины (Corylus), бука (Fagus) и белого клена (Acer pseudoplatanus).
Двудомные растения: мужские и женские половые органы образуются на разных растениях, т.е. каждое данное растение является либо мужским, либо женским, как, например, у тиса (Taxus), ивы (Salix), тополя (Populus) и остролиста (Ilex).
Гермафродитные (обоеполые) цветки: мужские и женские половые органы развиваются в одном и том же цветке, как у лютика, яснотки, пролески Endymion и гороха.
Однополые цветки: они несут либо только тычинки, либо только плодолистики, как, например, у дуба, лещины, тиса, тополя и остролиста.
Симметрия цветков
Если части цветка располагаются на цветоложе радиально-симметрично, то цветок называют правильным или актиноморфным (например, у лютика или пролески). Если же цветок обладает только двусторонней симметрией, то его называют неправильным или зигоморфным (например, у яснотки или гороха).
На рис. 20.15-20.18 изображено несколько типичных цветков, опыляемых насекомыми. Морфология цветка типичного злака — ветроопыляемого однодольного — представлена на рис. 3.38.

Рис. 20.15. Схематический продольный разрез цветка лютика (Ranunculus acris), отдел Spermatophyta, класс Angiospermae, подкласс Dicotyledonae, семейство Ranunculaceae). Это многолетнее травянистое растение, часто встречающееся на сырых лугах и пастбищах. Зимует благодаря наличию корневища. Цветки актиноморфные, свободнолепестные, с верхней завязью (подпестичные). Цветет с апреля по сентябрь. Опылители — двукрылые и мелкие перепончатокрылые. Каждый плодолистик содержит одно семя и образует плод, называемый семянкой. Специальных механизмов, способствующих распространению семян, нет

Рис. 20.16. Строение цветка душистого горошка (Lathyrus odoratus), отдел Spermatophyta, подкласс Dicotyledonae, семейство Leguminosae. Цветок актиноморфный, свободнолепестный. Цветет в июле. Чашечка состоит из пяти чашелистиков. Венчик пятилепестный: парус, два крыла и примыкающая к ним лодочка, образованная двумя сросшимися нижними лепестками. Венчик белый или окрашенный. Опыляется пчелами, которых привлекает окраска, запах и нектар. Особенно хорошо заметен парус. Крылья служат платформой, на которую садится насекомое. Под тяжестью его тела крылья опускаются вниз вместе с лодочкой, с которой они соединены. Столбик и рыльце выступают при этом наружу и соприкасаются с нижней поверхностью тела пчелы, на которой может находиться пыльца, захваченная с другого цветка. Пока пчела с помощью своего длинного хоботка добывает нектар у основания завязи, пыльца из пыльников наносится на нижнюю поверхность столбика, откуда она может попасть на пчелу. Иногда происходит и самоопыление. К душистому горошку близок горох огородный (Pisum sativum), но у него самоопыление более обычно. Плод — боб, состоящий из одного плодолистика со многими семенами. (Способ распространения семян — см. рис. 20.28.)

Рис. 20.17. Схематический продольный разрез цветка яснотки (Lamium album), отдел Spermatophyta, класс Angiospermae, подкласс Dicotyledonae, семейство Labiatae. Травянистое многолетнее растение, часто встречающееся у изгородей и на заброшенных землях. Перезимовывает благодаря наличию корневища. Цветки сростнолепестные, зигоморфные, двусторонне — симметричные, подпестичные. Цветет с апреля по июнь и осенью. Опыление происходит главным образом с помощью шмелей. Шмель садится на нижнюю губу венчика, и когда он проникает в цветок, его спинка, на которой может находиться пыльца, соприкасается с рыльцем, что приводит к перекрестному опылению, хотя у яснотки возможно и самоопыление. Плод — карцерула, состоящая из четырех орешков; каждый из них имеет твердую стенку и содержит по одному семени

Рис. 20.18. Схематический продольный разрез цветка пролески Endymion non-scripta, отдел Spermatophyta, класс Angiospermae, подкласс Monocotyledonae, семейство Liliaceae. Травянистое многолетнее растение, часто встречающееся в лесах и живых изгородях; предпочитает затененные участки и легкие кислые почвы. Перезимовывает при помощи клубней. Цветок актиноморфный, подпестичный. Число частей цветка кратно трем, что типично для однодольных. Цветет с апреля по май. Опыление производят насекомые, такие как медоносная пчела. Цветки издают слабый аромат; верхняя часть завязи выделяет нектар. Плод — коробочка, растрескивающаяся с образованием трех вальв
Строение цветка удобнее всего показать, если разрезать его по вертикали на две равные половинки (в медиальной плоскости, т. е. так, чтобы расщепить цветоножку надвое в продольном направлении) и изобразить одну половинку, очертив поверхность разреза непрерывной линией.
Развитие пыльцевых зерен
Каждая тычинка состоит из пыльника, который содержит четыре пыльцевых мешка, производящих пыльцу, и тычиночной нити, содержащей проводящий пучок, по которому в пыльник поступают питательные вещества и вода. На рис. 20.19 показано внутреннее строение пыльника с его четырьмя пыльцевыми мешками, содержащими микроспороциты, или материнские клетки микроспор. Каждый микроспороцит претерпевает мейоз и образует четыре пыльцевых зерна (рис. 20.20).

Рис. 20.19. Зрелый пыльник колокольчика в поперечном разрезе до и после растрескивания
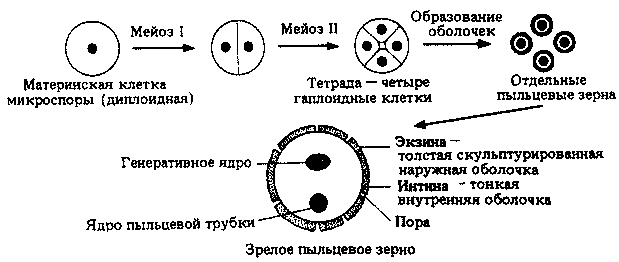
Рис. 20.20. Развитие пальцевых зерен
Непосредственно после мейоза можно видеть тетрады (группы по четыре) молодых пыльцевых зерен. У каждого пыльцевого зерна образуется толстая стенка, поверхность которой часто бывает причудливо скульптурирована, причем характер узоров специфичен для вида или рода. Наружная стенка, или экзина, состоит из спорополленина — материала, близкого к кутину и суберину, но более стойкого, чем оба этих вещества. Спорополленин — одно из самых устойчивых веществ, существующих в природе, и благодаря этому оболочки пыльцевых зерен могут сохраняться, не изменяясь, на протяжении длительного времени, иногда миллионов лет. Это обстоятельство, а также возможность довольно легко определять род или вид растения, которому принадлежало пыльцевое зерно, привели к созданию целой науки-палинологии, или пыльцевого анализа. Изучая пыльцевые зерна, относящиеся к определенному времени и собранные в определенном месте, можно установить, какие там росли растения, и получить информацию о существовавших в этом месте экосистемах (включая и животных) и климате. Особенно обильным источником пыльцевых зерен служит торф, мощные слои которого накапливаются на протяжении длительных периодов времени в торфяных болотах.
20.2. Как можно использовать пыльцевые зерна для определения а) климата данной области в прошлом и б) деятельности человека в те времена?
На этой стадии пыльцевое зерно эквивалентно микроспоре (см. гл. 3). Его ядро делится надвое путем митоза, образуя генеративное ядро и ядро пыльцевой трубки (рис. 20.20). После этого содержимое пыльцевого зерна можно рассматривать как эквивалент мужского гаметофита, поскольку из генеративного ядра в дальнейшем образуются мужские гаметы.
20.2.3. Развитие зародышевого мешка и женской гаметы
Каждый плодолистик состоит из рыльца, столбика и завязи. В завязи образуются один или несколько семязачатков; каждый из них прикреплен к стенке завязи в месте, называемом плацентой, с помощью короткой ножки-фуникулуса, по которому к развивающемуся семязачатку поступают питательные вещества и вода. Часто семязачаток в процессе своего развития загибается вниз, так что его кончик поворачивается к основанию фуникулуса; в таких случаях семязачаток называют анатропным. Если же фуникулус и семязачаток остаются прямыми, то семязачаток называют ортотропным.
Главную часть семязачатка составляет нуцеллус, окруженный двумя защитными покровами — интегументами. На одном конце семязачатка слева имеется маленькая пора — микропиле. Другой его конец, где фуникулус сливается с нуцеллусом и интегументами, называется халазой.
В нуцеллусе, у его микропилярного конца, начинает развиваться одна материнская клетка мегаспоры, известная под названием материнской клетки зародышевого мешка. Эта диплоидная клетка делится путем мейоза и образует гаплоидную мегаспору, или зародышевый мешок (рис. 20.21). Зародышевый мешок растет, его ядро делится путем митоза, и теперь его содержимое можно рассматривать как женский гаметофит. В результате дальнейших митозов образуется восемь ядер; одно из них — ядро женской гаметы.
Два полярных ядра перемещаются к центру зародышевого мешка и сливаются, превращаясь в одно диплоидное ядро (в некоторых случаях их слияние происходит позднее, при оплодотворении). Остальные шесть ядер, по три на каждом конце, разделяются тонкими клеточными стенками; по-видимому, лишь одно из них — ядро женской гаметы — принимает участие в дальнейшем развитии.

Рис. 20.21. Развитие зародышевого мешка и женской гаметы (в анатропном семязачатке)
Окончательный вид зрелого плодолистика перед оплодотворением показан на рис. 20.22.
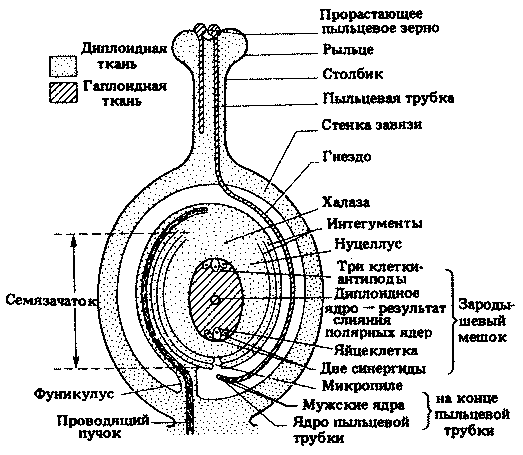
Рис. 20.22. Плодолистик в момент оплодотворения (в продольном разрезе). Обратите внимание, что семязачаток (анатропный), который после оплодотворения превращается в семя, содержит как диплоидную родительскую ткань, так и гаплоидную ткань зародышевого мешка. Содержимое пыльцевого зерна и пыльцевой трубки эквивалентно мужскому гаметофиту, а зародышевый мешок эквивалентен женскому гаметофиту
20.2.4. Опыление
После того как в пыльцевых мешках сформируются пыльцевые зерна, клетки, образующие стенки пыльника, начинают подсыхать и сморщиваться; возникает напряжение, которое в конце концов приводит к растрескиванию (раскрыванию) пыльников продольными щелями, образующимися на боковых поверхностях (рис. 20.19), что ведет к освобождению пыльцы.
Перенос пыльцевых зерен из пыльника на рыльце называется опылением. Опыление необходимо для того, чтобы мужские гаметы, развивающиеся в пыльцевых зернах, могли встретиться с женскими гаметами; в процессе эволюции возникли особые, нередко очень замысловатые, механизмы, обеспечивающие успешное опыление.
Перенос пыльцы с пыльника на рыльце того же самого цветка или другого цветка того же экземпляра растения называют самоопылением. Перенос пыльцы с пыльника одного экземпляра растения на рыльце другого называют перекрестным опылением. Перекрестное опыление ведет к перекрестному оплодотворению и способствует генетической изменчивости. Таким образом, это особого рода аутбридинг. У многих растений имеются специальные особенности, благоприятствующие перекрестному опылению; некоторые из них будут описаны ниже.
Преимущество самоопыления, ведущего к самооплодотворению, состоит в том, что оно более надежно, особенно в тех случаях, когда представители данного вида встречаются относительно редко и на больших расстояниях один от другого. Это связано с тем, что самоопыление не зависит от внешних агентов, таких как ветер или насекомые. Однако самооплодотворение, будучи крайней формой инбридинга, может приводить к уменьшению жизнеспособности потомков (см. разд. 25.4). Примерами самоопыляющихся растений могут служить крестовник (Senecio) и звездчатка (Stellaria); их цветки не образуют нектара и лишены запаха.
Как перекрестное опыление, так и самоопыление имеют свои преимущества и свои недостатки, и у многих растений выработались приспособления, которые, благоприятствуя перекрестному опылению, вместе с тем дают возможность, если оно почему-либо не удалось, прибегнуть к самооплодотворению. Например, у фиалки (Viola) и у кислицы (Oxalis) некоторые бутоны не раскрываются, так что для них неизбежно самоопыление.
Особенности, благоприятствующие самоопылению
Двудомные растения. У двудомных растений самоопыление невозможно. Однодомные виды, у которых на одном и том же гермафродитном растении есть и мужские, и женские цветки, тоже предпочитают перекрестное опыление, но у них может проис-ходить и самоопыление.
20.3. Двудомные растения встречаются редко, несмотря на преимущества перекрестного опыления, Приведите две возможные причины этого.
20.4. "Двудомность" (раздельнополость) широко распространена у животных. Почему эта система оказалась более подходящей для животных, чем для цветковых растений?
Дихогамия. Иногда созревание пыльников и рылец происходит в разные сроки — явление, называемое дихогамией. Если пыльники созревают раньше, чем рыльца, то говорят о протандрии, а если сначала созревают рыльца — о протогинии. Протандрия встречается гораздо чаще; примерами служат яснотка, одуванчик, иван-чай (Epilobium angustifolium), а также шалфей (Salvia, рис. 20.23).
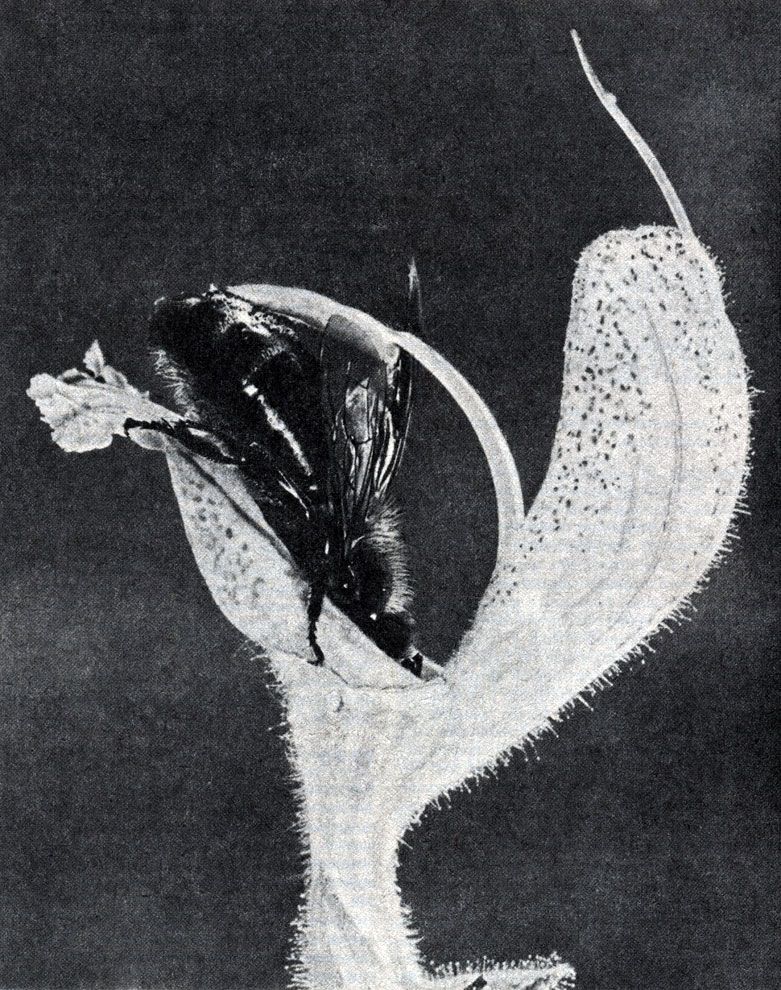
Рис. 20.23. Пчела, входящая в цветок лугового шалфея. Голова пчелы надавливает на пластинку, к которой прикреплены тычинки, и они опускаются, стряхивая пыльцу на брюшко пчелы. Если пчела посетит теперь более старый цветок (по мере старения цветка его рыльце удлиняется), то ее брюшко соприкоснется с рыльцем и приставшая к брюшку пыльца будет перенесена на рыльце. В результате всего этого произойдет перекрестное опыление
Протогиния свойственна пролеске и норичнику (Scrophularia). При дихогамии в большинстве случаев имеется такой период, когда и пыльники, и рыльца находятся в состоянии зрелости, что делает возможным самоопыление, если перекрестное опыление не произошло. Сходный механизм, обеспечивающий перекрестное оплодотворение, существует у некоторых гермафродитных животных, например у гидры, у которой мужские гонады созревают раньше, чем женские.
Самонесовместимость (самостерильность). Даже если самоопыление произошло, пыльцевые зерна часто не развиваются или развиваются очень медленно, что препятствует самооплодотворению или уменьшает его вероятность. Во всех таких случаях происходит специфическое взаимодействие, ингибирующее процесс прорастания пыльцевой трубки в столбик, причем это ингибирование определяется генами самонесовместимости.
В случаях самонесовместимости частота совместимых перекрестных комбинаций варьирует и опять-таки генетически детерминирована. Для максимально эффективного использования пыльцы значительная доля скрещиваний должна быть совместимой. Крайним примером служит клевер, у которого все растения самонесовместимы, но перекрестная несовместимость встречается менее чем в одной из 22000 пар. Менее эффективны системы, в которых совместимые типы различаются по морфологии цветков. Примером служит первоцвет (см. ниже).
20.5. Самонесовместимость контролируется множественными аллелями. Если принять, что а) имеется три аллеля — S1, S2 и S3 и б) самонесовместимость возникает в тех случаях, когда пыльцевое зерно и клетки рыльца имеют один общий аллель, то какая доля пыльцевых зерен от растения с генотипом S1 S2 будет способна успешно прорасти на растении с генотипом S2 S3?
Структурные адаптации. У большинства гермафродитных цветков имеются структурные особенности, способствующие перекрестному опылению.
У энтомофильных (опыляемых насекомыми) цветков рыльца обычно выступают над пыльниками, что не позволяет пыльцевым зернам падать прямо на рыльца того же цветка. Когда насекомое, несущее на себе пыльцу с другого растения, посещает такой цветок, оно сначала прикасается к его рыльцу. Затем, когда насекомое ищет нектар, оно либо обмазывается пыльцой, либо стряхивает ее на себя. Так обстоит дело, например, при опылении яснотки (рис. 20.17). Более примитивный механизм состоит в том, что рыльце прикасается к насекомому, когда оно садится на цветок, как при опылении душистого горошка (см. рис. 20.16). Такие механизмы обычно сочетаются с дихогамией, и цветки при этом часто бывают сложными и зигоморфными.
Цветки привлекают насекомых, предоставляя им пищу (нектар или пыльцу) и стимулируя их зрение и обоняние. Это возможно благодаря наличию у цветков особых признаков, которые будут рассмотрены ниже.
У многих ветроопыляемых цветков тычинки, цветок в целом или все соцветие свисают вниз, так что пыльца высыпается, а затем уносится прочь (как, например, у лещины).
20.6. На рис. 20.24 изображены цветки первоцвета двух типов, встречающихся в природе примерно в равном количестве и различающихся длиной столбиков (гетеростилия) и расположением тычинок, а) Известно, что пчелы высасывают нектар из нижней части трубки венчика; объясните, почему перекрестное опыление происходит главным образом между длинно- и короткостолбчатыми цветками, а не между цветками одного типа? б) В чем преимущество такой системы?
Хотя гетеростилия (см. вопрос 20.6 и рис. 20.24), очевидно, способствует аутбридингу, гораздо более эффективен в этом смысле механизм самонесовместимости, существующий у длинно- и короткостолбчатых первоцветов, благодаря которому перекрестное оплодотворение происходит только между цветками разных типов. Гены, контролирующие несовместимость, длину столбика и высоту расположения тычинок, локализованы очень близко друг к другу в одной и той же хромосоме и ведут себя как одна наследуемая единица.

Рис. 20.24. Гетеростилия у первоцвета (Primula). А. Длинностолбчатый цветок. Б. Короткостолбчатый цветок
Опыление ветром и опыление насекомыми
Пыльцевые зерна — это споры, но в отличие от спор, образуемых споровыми растениями, они не могут прорастать на суше и должны быть перенесены на женские половые структуры шишек (у голосеменных) или цветков (у покрытосеменных). Первоначально споры распространялись ветром, но такой механизм переноса пыльцы очень неэффективен, так как при этом попадание пыльцевых зерен на шишки или цветки всецело зависит от случая. Все хвойные и многие цветковые растения (злаки и большинство деревьев умеренного пояса, такие как дуб и лещина) по-прежнему опыляются с помощью ветра, но им приходится производить огромные количества пыльцы, что требует больших затрат вещества и энергии. После того как в процессе эволюции возникли цветки, растения очень быстро начали использовать для опыления насекомых, так как это гораздо более надежный способ переноса пыльцы. Насекомое может переносить небольшое количество пыльцы с тычинок одного цветка точно на рыльце другого. В результате в процессе эволюции между цветками и насекомыми сложились особые взаимоотношения: наградой, которую насекомые получают от цветков, служит пища в виде нектара, а иногда пыльцы. Насекомые, специализированные для питания на цветках, появились одновременно с цветковыми растениями; к их числу относятся пчелы, шмели, осы и бабочки. В некоторых случаях насекомое и опыляемое им растение настолько взаимозависимы, что ни одно из них не может существовать без другого, как, например, юкка и юкковая моль (Pronuba yuccasella). Энтомофилия обладает еще одним преимуществом: она благоприпятствует перекрестному опылению, а тем самым и перекрестному оплодотворению; поэтому описанные ниже модификации цветков, облегчающие опыление насекомыми, можно добавить к перечню признаков, способствующих перекрестному опылению.
У энтомофильных растений цветки обычно крупные, с ярко окрашенными лепестками, что привлекает насекомых; если же цветки мелкие, то они собраны в соцветия. Насекомые воспринимают лучи ультрафиолетовой области спектра, невидимые для человека, а поэтому цветки, кажущиеся человеку белыми, насекомые могут воспринимать как окрашенные. Часто на лепестках имеются полоски, пятнышки или более интенсивно окрашенные участки, указывающие насекомым путь к нектарникам; они есть, например, у фиалок, включая анютины глазки (Viola), орхидей (Orchis и другие роды) и наперстянки (Digitalis).
Более специфичны, чем окраска, запахи, издаваемые цветками; некоторые из них, например лаванда и роза, используются в парфюмерной промышленности. Некоторые растения обладают запахом гниющего мяса, который привлекает насекомых, питающихся падалью; арум (Arum maculatum) привлекает навозных мух. Специфическим опознавательным признаком может служить также форма цветка.
Один из самых сложных и странных механизмов для обеспечения перекрестного опыления обнаружен у некоторых орхидей, цветки которых по форме, окраске и даже запаху подражают самкам ос — сфексов и притом настолько убедительно, что сфексы — самцы пытаются копулировать с цветками (рис. 20.25). При этих попытках насекомое стряхивает с себя пыльцу на цветок, а затем, покидая его, уносит на себе его пыльцу, которую передает потом другому цветку.
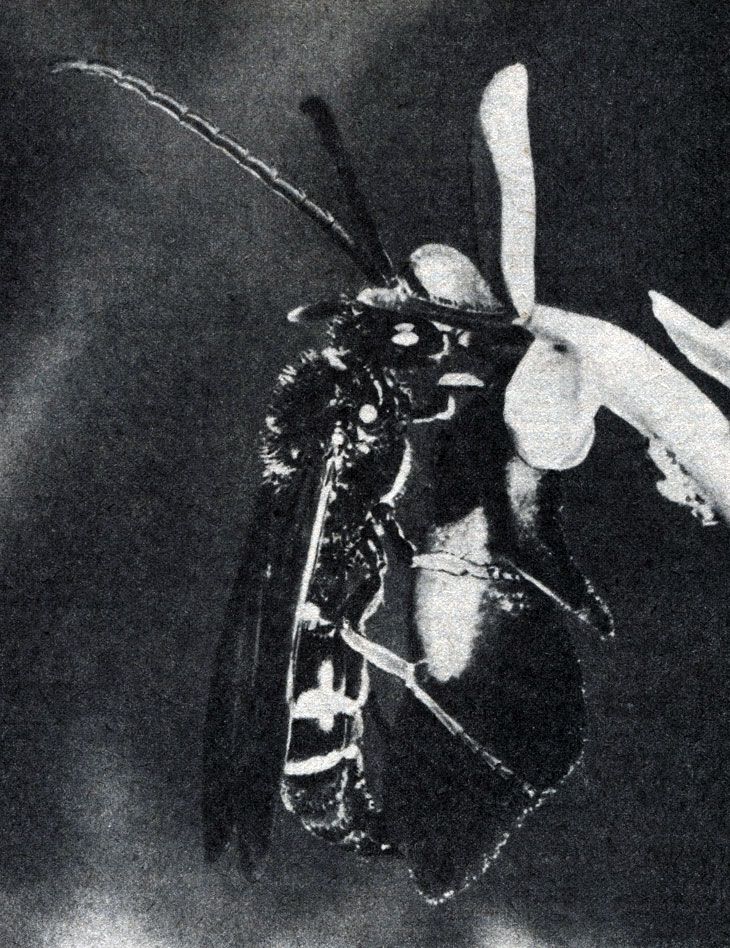
Рис. 20.25. Роющая оса Argogorytes mystaceus, пытающаяся копулировать с цветком офриса пчелоносного (Ophrys insectifera)
Типичные различия между цветками, опыляемыми ветром и опыляемыми насекомыми, приведены в табл. 20.2.

Таблица 20.2. Типичные различия между цветками, опыляемыми ветром и цветками, опыляемыми насекомыми
20.2.5. Оплодотворение
Как только на рыльце попадает пыльцевое зерно, эпидермальные клетки рыльца выделяют раствор сахарозы, который стимулирует прорастание пыльцевого зерна и, возможно, используется для его питания (рис. 20.26). Сквозь одну из пор, имеющихся в стенке пыльцевого зерна, выходит пыльцевая трубка, которая быстро растет вниз внутри столбика, направляясь к завязи. Ее рост сопровождается секрецией пищеварительных ферментов и регулируется полученным от пыльцевого зерна ядром пыльцевой трубки, которое находится у ее растущего кончика. Рост пыльцевой трубки стимулируют ауксины, вырабатываемые гинецеем, а к завязи ее направляют определенные вещества (пример хемотропизма). Возможно также, что здесь имеет место отрицательный аэротропизм, т. е. рост в направлении от воздушной среды. Как уже говорилось, рост зависит от совместимости между пыльцой и тканью рыльца.

Рис. 20.26. Микрофотогрария пыльцевых зерен на рыльце цветка, сделанная с помощью сканирующего электронного микроскопа Шиповатая поверхность пыльцевых зерен характерна для цветков опыляемых насекомыми
Во время роста пыльцевой трубки генеративное ядро пыльцевого зерна делится митотически, образуя два мужских ядра, представляющие собой мужские гаметы (рис. 20.22). В отличие от спермиев низших растений они неподвижны и могут добраться до женской гаметы, находящейся в зародышевом мешке семязачатка, только с помощью пыльцевой трубки. Пыльцевая трубка проникает в семязачаток через микропиле, ее ядро дегенерирует, а кончик трубки разрывается, освобождая мужские гаметы вблизи зародышевого мешка, в который они и проникают. Одно ядро сливается с женской гаметой, образуя диплоидную зиготу, а другое — с двумя полярными ядрами (или с диплоидным ядром, если эти последние уже слились), образуя триплоидное ядро, называемое первичным ядром эндосперма. Такое двойное оплодотворение свойственно только цветковым растениям.
Если, как это часто бывает, гинецей содержит несколько семязачатков, то каждый из них, чтобы превратиться в семя, должен быть оплодотворен отдельным пыльцевым зерном. Поэтому может оказаться, что один цветок даст семена, образовавшиеся в результате оплодотворения пыльцевыми зернами от разных растений.
Опыт 20.1. Изучение роста пыльцевых трубок
Рыльца выделяют раствор, содержащий от 2 до 45% сахарозы. Этот раствор способствует прилипанию пыльцевых зерен к рыльцу и их прорастанию. Добавив к экспериментальному раствору борат натрия, можно предотвратить осмотический разрыв кончиков пыльцевых трубок и стимулировать рост.
Микроскоп
Предметное стекло с лункой
Цветки с растрескивающимися пыльниками, например цветки яснотки или лакфиоли 10-20%-ный (масса/объем) раствор сахарозы, содержащий также борат натрия (0,01%)
Ацетокармин или нейтральный красный
Каплю раствора сахарозы помещают в лунку предметного стекла и вносят в нее пыльцевые зерна, прикасаясь к капле поверхностью растрескавшегося пыльника. Препарат рассматривают под микроскопом спустя различные промежутки времени на протяжении 1-2 часов. Ядра, находящиеся в кончике растущих пыльцевых трубок, можно окрасить, добавив к раствору каплю ацетокармина или нейтрального красного.
20.2.6. Развитие семени вплоть до стадии покоя
После оплодотворения семязачаток называют уже семенем, а завязь — плодом.
Зигота растет путем многократных митотических делений и превращается в многоклеточный зародыш, состоящий из первичного побега, или плюмулы, первичного корешка и одной (у однодольных) или двух (у двудольных) семядолей. Семядоли имеют более простую структуру, чем первые настоящие листья; иногда они наполнены питательными веществами и служат запасающей тканью, как, например, у гороха и у конских бобов (Vicia faba). Плюмула состоит из стебля, первой пары настоящих листьев и терминальной почки. Триплоидное ядро первичного эндосперма многократно делится (путем митоза), образуя эндосперм — скопление триплоидных ядер, отделенных друг от друга тонкими клеточными стенками. У некоторых растений, например у кукурузы (Zea mays), эндосперм сохраняет роль запасающей ткани.
Если запасающей тканью служат семядоли, то они растут за счет эндосперма, который может при этом совершенно исчезнуть. У некоторых семян запасы питательных веществ содержатся и в эндосперме, и в семядолях.
В развивающемся семени рост зародыша, а иногда и эндосперма происходит в зародышевом мешке. По мере продолжения роста окружающий их нуцеллус дезорганизуется и разрушается, доставляя необходимые для роста питательные вещества. В дальнейшем снабжение питательными веществами обеспечивает проводящий пучок фуникулуса (ножки семязачатка), подобно тому как развивающийся зародыш млекопитающего получает питание от материнского организма через плаценту. Как уже отмечалось, место прикрепления фуникулуса к стенке завязи тоже называется плацентой.
Микропиле сохраняется в виде маленькой поры в тесте (семенной кожуре), через которую поступают кислород и вода при прорастании семени. Теста — это тонкий, но прочный защитный слой, образующийся из интегументов. На завершающих стадиях созревания семени содержание в нем воды снижается от обычного для растительных тканей — примерно 90% (по массе) — до 10-15%. Это сопровождается заметным снижением метаболической активности и представляет собой важный шаг к переходу семени в состояние покоя.
По мере развития семени завязь превращается в зрелый плод и стенка ее получает название перикарпия. Происходящие при этом изменения у разных видов различны, но обычно направлены на то, чтобы плод мог защищать семена и способствовать их распространению (см. разд. 20.2.8).
Остальные части цветка увядают, отмирают и сбрасываются, подобно листьям у листопадных растений. В некоторых случаях такие части, как цветоложе, столбик или чашелистики сохраняются и участвуют в распространении семян; цветоложе в таких случаях, например у земляники (Fragaria), получает название ложного плода.
Гормональная регуляция развития плода рассмотрена в разд. 15.3.5.
Некоторые изменения, происходящие после оплодотворения, суммированы в табл. 20.3.

Таблица 20.3. Изменения, происходящие у цветковых растений после оплодотворения
Некоторые стадии развития зародыша показаны на рис. 20.27.

Рис. 20.27. Развитие зародыша в лишенном эндосперма семени двудольного растения — пастушьей сумки (Capsella bursa-pastoris)
20.2.7. Преимущества и недостатки размножения семенами
Семя — типичный продукт полового размножения у семенных растений; согласно определению, это оплодотворенный семязачаток. Семя содержит зародыш растения и одну или более семядолей, а иногда эндосперм, и окружено защитной кожурой.
Преимущества
1. Растение не нуждается в водной среде для полового размножения и поэтому лучше приспособлено к наземным условиям.
2. Семя защищает зародыш.
3. Семя содержит питательные вещества, необходимые зародышу (либо в семядолях, либо в эндосперме).
4. Семя обычно имеет приспособления для распространения.
5. Семя способно оставаться в состоянии покоя и переживать неблагоприятные условия.
6. Семя реагирует на благоприятные условия и иногда должно пройти период дозревания, т.е. прорастает не сразу (см. гл. 15).
7. Семена образуются в результате полового размножения, а поэтому обеспечивают виду преимущества, связанные с генетической изменчивостью.
Недостатки
1. Семена имеют относительно крупные размеры из-за находящихся в них больших запасов питательных веществ. Это затрудняет их распространение по сравнению со спорами.
2. Животные часто поедают семена ради содержащихся в них питательных веществ.
3. Опыление часто зависит от внешних факторов (ветер, насекомые, вода). Поэтому оно (а значит, и оплодотворение) не всегда гарантировано, особенно если производится ветром.
4. Потери семян велики, так как вероятность выживания каждого данного семени незначительна. Поэтому для обеспечения успеха родительский спорофит должен вкладывать в производство семян большие количества вещества и энергии.
5. Запасы питательных веществ в семени ограниченны, тогда как при вегетативном размножении родительское растение доставляет дочернему растению питательные вещества до тех пор, пока это второе не станет вполне самостоятельным.
6. У двудомных видов в размножении участвуют два растения, что повышает вероятность неудачи по сравнению с размножением при участии одной родительской особи. Напомним, что двудомность-явление относительно редкое. Приведенные выше сведения можно использовать для сопоставления выгодных или невыгодных сторон организации семенных растений по сравнению со споровыми или относительных преимуществ полового и вегетативного размножения у семенных растений.
20.2.8. Распространение плодов и семян
После образования семян либо весь плод, либо содержащиеся в нем семена (или семя) отделяются от родительского спорофита. Если перикарпий (стенка плода) становится твердым и сухим, плод называют сухим, если же он становится мясистым, то плод называют сочным. Сухой плод может быть вскрывающимся или невскрывающимся в зависимости от того, связано ли освобождение семян с разрывом его стенки или нет. Сухие плоды могут также распадаться на несколько односеменных члеников; такие плоды называют схизокарпными.
Чем дальше разносятся семена, тем менее вероятна конкуренция со стороны родительского растения. Это дает также больше шансов колонизировать новую территорию, что со временем приводит к увеличению размеров популяции в целом. Однако при этом существует опасность, что семя, будучи занесено слишком далеко от родительского растения, не найдет подходящего места для прорастания.
Существуют три главных внешних фактора, распространяющих семена, — ветер, животные и вода. Кроме того, имеются механизмы автохории, часто связанные со взрывным освобождением семян из плода. Примеры всех этих способов распространения семян представлены на рис. 20.28.
Распространение животными
Распространение семян животными в общем более надежно, так как животные обычно посещают плодородные участки, подходящие для прорастания семян. Плоды, снабженные колючками или крючочками, прицепляются к коже или шерсти проходящих мимо животных и могут быть перенесены на некоторое расстояние, прежде чем отпадут или будут содраны. Прицепки развиваются из разных частей плода или окружающих структур. Примерами служат плоды подмаренника цепкого (Galium aparine), гравилата (Geum), лопуха (Arctium), моркови (Daucus), лютика (Ranunculus), череды (Bidens), репешка (Agrimonia). У гравилата крючочки имеются на столбике. У лопуха скопление мелких плодов окружено крючковидными листочками обертки (видоизмененные листья), образующими единую структуру; кроме того, здесь имеются мелкие жесткие волоски, которые проникают в кожу, вызывая раздражение и расчесывание, в результате чего плоды отпадают. У подмаренника, моркови и лютика перикарпий покрыт выростами (прицепками). У череды плод снабжен летучкой, как у одуванчика, но с прочными шипами. Цветоложе репешка покрыто жесткими загнутыми колючками.

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Подмаренника цепкого (Galium aparine) и репешок

Плод подмаренника с многочисленными крючочками, развивающимися из околоплодника. Микрофотография, полученная с помощью сканирующего электронного микроскопа.
Примерами сочных плодов служат плоды сливы (Prunus), ежевики (Rubus), томата (Lycopersicum), яблони (Malus), земляники (Fragaria). Мясистые части этих плодов служат пищей животным, в том числе птицам. При созревании они обычно приобретают яркую окраску и аромат, привлекающие животных. Животное съедает и переваривает плод, но семена, устойчивые к пищеварительным ферментам, проходят невредимыми через его кишечник и выходят с испражнениями, часто попадая на плодородную почву. Содержащиеся в экскрементах питательные вещества могут повышать плодородие почвы вокруг семени.
Слива
Слива служит примером плода, называемого костянкой; ее перикарпий состоит из трех слоев: экзокарпия (защитной кожицы), сочного мезокарпия и внутреннего каменистого эндокарпия ("косточки"). Эти слои защищают семя, предохраняя его от переваривания. Другие примеры костянок — плоды персика, вишни и миндаля (также принадлежащих к роду Prunus), бузины (Sambucus) и кокосовой пальмы (Cocos nucifera; плод с волокнистым мезокарпием). Миндаль, поступающий в продажу, обычно очищают от зкзо- и мезокарпия.

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Слива (в продольном разрезе)

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Ягода ежевики в продольном разрезе — совокупность костянок, образовавшихся из одного цветка
Томат
Плод томата, образующийся в результате слияния двух, трех или четырех плодолистиков, относится к типу ягодовидных. Он сходен с костянкой, но эндокарпий у него мясистый, а не каменистый. К ягодовидным принадлежат также плоды черной смородины и крыжовника (Ribes spp.), тыквы (Cucurbita), апельсина и лимона (Citrus spp.), банана (Musa), винограда (Vitis), финика (Phoenix dactylifera; содержит одно деревянистое семя).

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Томат (в продольном разрезе)
Иногда цветоложе разбухает, становится мясистым и само напоминает плод. Такие плоды называют ложными. Примером служат плоды яблони и земляники.
Яблоко
Ложный плод яблони относится к типу яблока, или тыквины. У плодов этого типа мякоть образуется из полого цветоложа, окружающего заключенные в нем плодолистики. Перикарпий настоящего плода превращается в "сердцевину", содержащую семена. Другими примерами служит груша (Pyrus) и боярышник (Crataegus)

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Яблоко (в продольном разрезе)
Земляника
У земляники мелкие зеленые плоды, называемые семянками и расположенные на разросшемся цветоложе; каждая семянка содержит по одному семени.
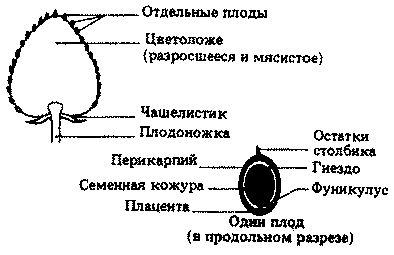
Рис. 20.28. Разные типы плодов и семян и способы их распространения. Ягода земляники (в продольном разрезе)
Ореховидные плоды, например у дуба (Quercus), бука (Fagus), каштана ( Casta nеа), относительно крупные сухие плоды, которые не расщепляются и не выбрасывают семена. Распространяются ореховидные плоды целиком, часто при помощи животных, делающих запасы, в особенности таких грызунов, как белки. На рисунке изображен плод дуба, называемый желудем.

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Плод дуба — желудь
Распространение ветром
Многие растения, распространяемые ветром, имеют специальные приспособления. К их числу относятся летучки, которыми снабжены, например, семена кипрея (Epilobium), ивы (Salix), хлопчатника (Gossypium hirsutum), плоды одуванчика (Taraxacum), ломоноса (Clematis), и крылатки, как, например, у сосны (голосеменные), вяза (Ulmus), ясеня (Fraxinus), клена (Acer), граба (Carpinus).

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Одуванчик, клен (Acer pseudoplatanum), кипрей
Плод клена до или после того, как он будет перенесен ветром на новое место, распадается на две части по шву между плодолистиками (такой плод называют схизокарпным); опадающий плод вращается и подхватывается ветром.
У ряда растений, например у мака (Paraver), чернушки (Nigella), наперстянки (Digitalis), лихниса (Lichnis), плод — коробочка, сидящая на конце длинного стебелька, которую колышет ветер, так что многочисленные мелкие семена высыпаются через поры в верхней части плода.

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Мак и коробочка (в продольнрм разрезе)
Автохория
Семена некоторых растений, например орхидей, настолько мелкие и легкие, что разносятся ветром, не имея никаких специальных придатков, которые бы увеличивали их поверхность. Такое распространение, называемое обычно автохорией, наблюдается у вскрывающихся плодов. Вскрывание у них часто происходит путем растрескивания по шву между плодолистиками, как, например, у бобов. Семена при этом либо выбрасываются из плодов более или менее сильным толчком, либо просто выпадают из них. Некоторые из таких семян имеют приспособления для распространения ветром (например, у кипрея). Коробочки тоже бывают вскрывающимися (например, у мака и кипрея), причем у некоторых растений семена выбрасываются из них с силой (например, у фиалки).
Бобы — плоды, характерные для представителей сем. бобовых (Leguminosae), например для гороха (Pisum), конских бобов (Vicia), фасоли (Phaseolus), утесника (Ulex), ракитника (Cytisus), золотого дождя (Laburnum). Боб состоит из одного плодолистика, содержащего много семян. Перикарпий, высыхая, скручивается, так как его наружные и внутренние волокнистые слои сокращаются в разных косых направлениях; в результате создается напряжение, и боб может внезапно раскрыться по двум боковым швам, с силой выталкивая наружу несколько семян. Другие семена выбрасываются менее энергично по мере дальнейшего скручивания (ранние стадии развития плода гороха показаны на рис. 20.16).

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Горох или ракитник
Распространение водой
Лишь немногие плоды и семена специально приспособлены для распространения с помощью воды. Они содержат воздушные полости, удерживающие их на поверхности воды. Кокосовый орех (плод Cocos nucifera) — костянка (ср. со сливой) с волокнистым мезокарпием и многочисленными воздухоносными полостями. У кувшинки (Nymphaea) семя снабжено губчатой оболочкой (ариллусом), происходящей из ножки семязачатка.

Рис. 20.28. Разные типы плодов и семян и способы их распространения. Кувшинка
Случайное распространение
Разделение семян и плодов на категории в зависимости от способа распространения никак нельзя считать строгим, и степень специализации к распространению ветром, животными или автохории сильно варьирует. Очевидно, во многих случаях большую роль играет элемент случайности, и данный плод или семя может распространяться двумя из названных трех способов или даже всеми тремя. Один из главных факторов случайного распространения является человек; семена могут прицепляться или прилипать к его одежде и т. п. или перевозиться вместе с разными грузами на транспортных средствах. Засорение зерновых культур семенами сорняков — обычное явление, наблюдаемое по всему земному шару. Орехи, припрятанные про запас грызунами, могут остаться и прорасти следующей весной. Наводнения, ураганы и т. п. могут занести семена дальше, чем обычно.
20.3. Половое размножение у человека
20.3.1. Мужская половая система
Мужская половая система состоит из парных семенников (яичек), семявыносящих протоков, ряда придаточных желез и полового члена (пениса). Семенник — сложная трубчатая железа яйцевидной формы; она заключена в капсулу — белочную оболочку — и состоит примерно из тысячи сильно извитых семенных канальцев, погруженных в соединительную ткань, в которой содержатся интерстициальные (лейдиговы) клетки. В семенных канальцах образуются гаметы — спермии (сперматозоиды), а интерстициальные клетки вырабатывают мужской половой гормон тестостерон. Семенники расположены вне брюшной полости, в мошонке, а поэтому спермии развиваются при температуре, которая на 2-3 °С ниже температуры внутренних областей тела. Более низкая температура мошонки частично определяется ее положением, а частично-сосудистым сплетением, образуемым артерией и веной семенника и действующим как противоточный теплообменник. Сокращения особых мышц перемещают семенники ближе или дальше от тела в зависимости от температуры воздуха, чтобы поддерживать температуру в мошонке на уровне, оптимальном для образования спермы. Если мужчина достиг половой зрелости, а семенники не опустились в мошонку (состояние, называемое крипторхизмом), то он навсегда остается стерильным, а у мужчин, носящих слишком тесные трусы или принимающих очень горячие ванны, образование спермиев может так сильно понизиться, что это приведет к бесплодию. Лишь у немногих млекопитающих, в том числе у китов и слонов, семенники всю жизнь находятся в брюшной полости.
Семенные канальцы достигают 50 см в длину и 200 мкм в диаметре и расположены в участках, называемых дольками семенника. Оба конца канальцев соединяются с центральной областью семенника — сетью семенника (rete testis) — короткими прямыми семенными канальцами. Здесь сперма собирается в 10-20 выносящих канальцах; по ним она переносится в головку придатка (эпидидимиса), где концентрируется в результате обратного всасывания жидкости, выделяемой семенными канальцами. В головке придатка спермии созревают, после чего они проходят по извитому 5-метровому выносящему канальцу к основанию придатка; здесь они остаются в течение короткого времени, прежде чем попадают в семявыносящий проток. Семявыносящий проток — это прямая трубка длиной около 40 см, которая вместе с артерией и веной семенника образует семенной канатик и переносит сперму в уретру (мочеиспускательный канал), проходящую внутри полового члена. Взаимоотношения между этими структурами, мужскими придаточными железами и половым членом показаны на рис. 20.29 и 20.30.
Рис. 20.29 Схема строения мужской половой системы

Рис. 20.30. Упрощенная схема строения семенника человека и протоков, по которым сперма поступает из семенных канальцев в уретру
Образование гамет (гаметогенез)
Клетки зачаткового эпителия в мужских и женских половых железах (гонадах) претерпевают ряд последовательных митотических и мейотических делений, называемых в совокупности гаметогенезом, в результате чего образуются зрелые мужские гаметы (сперматогенез) и женские гаметы (оогенез). В обоих случаях процесс делится на три фазы — фазу размножения, фазу роста и фазу созревания. Фаза размножения включает многократные митотические деления, приводящие к образованию множества сперматогоний или оогоний. Каждая из них проходит период роста в порядке подготовки к первому делению мейоза и последующему цитокинезу. Затем начинается фаза созревания, во время которой происходят первое и второе деления мейоза с последующей дифференцировкой гаплоидных клеток и формированием зрелых гамет.
Развитие сперматозоидов (сперматогенез)
Сперматозоиды (спермии) образуются в результате ряда последовательных клеточных делений, называемых в совокупности сперматогенезом, за которыми следует сложный процесс дифференцировки, называемый спермиогенезом (рис. 20.31). Процесс образования спермия занимает примерно 70 дней; на 1 г веса яичка образуется 107 спермиев в сутки. Эпителий семенного канальца состоит из наружного слоя клеток зачаткового эпителия и примерно шести слоев клеток, образовавшихся в результате многократных делений клеток этого слоя (рис. 20.32 и 20.33); эти слои соответствуют последовательным стадиям развития сперматозоидов. Сначала деление клеток зачаткового эпителия дает начало многочисленным сперматогониям, которые увеличиваются в размерах и становятся сперматоцитами первого порядка. Эти сперматоциты в результате первого деления мейоза образуют гаплоидные сперматоциты второго порядка, после чего претерпевают второе деление мейоза и превращаются в сперматиды. Между "тяжами" из развивающихся клеток расположены крупные клетки Сертоли, или трофические клетки, расположенные во всем пространстве от наружного слоя канальца до его просвета.

Рис. 20.31 Схема процессов сперматогенеза и спермиогенеза у человека

Рис. 20.32. Микрофотография среза семенника. Видны семенные канальцы и лейдиговы (интерстициальные) клетки

Рис. 20.33. Участок стенки семенного канальца (схема). Показаны лейдиговы клетки и различные стадии сперматогенеза и спермиогенеза
20.7. Клетки Сертоли богаты элементами гладкого эндоплазматического ретикулума, содержат обширный аппарат Гольджи и много митохондрий и лизосом. Что можно сказать о функции этих клеток на основании такой их структуры?
Сперматоциты располагаются в многочисленных впячиваниях на боковых поверхностях клеток Сертоли; здесь они превращаются в сперматиды, а затем переходят на тот край клетки Сертоли, который обращен к просвету семенного канальца, где они созревают, образуя спермии (рис. 20.33). По-видимому, клетки Сертоли обеспечивают созревающим спермиям механическую опору, защиту и питание. Все питательные вещества и кислород, доставляемые развивающимся гаметам по окружающим семенные канальцы кровеносным сосудам, и выделяемые в кровь отходы метаболизма проходят через клетки Сертоли. Эти клетки секретируют и жидкость, с которой спермии проходят по канальцам.
Сперматозоиды
Сперматозоиды, или спермии, — это очень мелкие подвижные мужские гаметы, образуемые мужскими гонадами-семенниками; их количество исчисляется миллионами. Форма спермиев у разных животных различна, однако строение их однотипно. Каждый сперматозоид можно подразделить на пять участков (рис. 20.34). В головке сперматозоида находится ядро, содержащее гаплоидное число хромосом и прикрытое акросомой. Акросома — особая структура, ограниченная мембраной, — содержит гидролитические ферменты, способствующие проникновению спермия в ооцит непосредственно перед оплодотворением; таким образом, функционально ее можно рассматривать как увеличенную лизосому. В короткой шейке спермия расположена пара центриолей, лежащих под прямым углом друг к другу. Микротрубочки одной из центриолей удлиняются, образуя осевую нить жгутика, проходящую вдоль всей остальной части сперматозоида. Средняя часть расширена за счет содержащихся в ней многочисленных митохондрий, собранных в спираль вокруг жгутика. Эти митохондрии доставляют энергию для сократительных механизмов, обеспечивающих движения жгутика. Главная и хвостовая части сперматозоида имеют строение, характерное для жгутиков: на поперечном срезе видна типичная структура "9 + 2" — 9 пар периферических микротрубочек, окружающих пару центральных микротрубочек.

Рис. 20.34. Схема строения зрелого спермия человека
Если смотреть на головку человеческого спермия сверху, то она кажется округлой, а при взгляде сбоку — уплощенной. Одного только жгутикового движения недостаточно, чтобы сперматозоид мог пройти расстояние от влагалища до места, где происходит оплодотворение. Главная локомоторная задача спермиев состоит в том, чтобы скопиться вокруг ооцита и ориентироваться определенным образом, прежде чем проникнуть сквозь мембраны ооцита.
Эндокринная функция яичек у человека
Рост, развитие и поддержание яичек у человека контролируются гонадотропными гормонами гипофиза — фолликулостимулирующим (ФСГ) и лютеинизирующим (ЛГ). ФСГ стимулирует развитие спермиев, а ЛГ-синтез стероидного гормона тестостерона лейдиговыми клетками яичка. ЛГ оказывает влияние на эти клетки, заставляя их синтезировать циклический аденозинмонофосфат (цАМФ); цАМФ переходит в ядро, где воздействует на генетический аппарат и таким образом стимулирует синтез ферментов, участвующих в образовании тестостерона из холестерола. (У мужчин ЛГ иногда называют по месту его действия гормоном, стимулирующим интерстициальные клетки).
Тестостерон — это главный андрогенный гормон, влияющий как на первичные, так и на вторичные половые признаки. Для успешного образования спермиев необходимы и тестостерон, и ФСГ, тогда как развитие вторичных половых признаков в период полового созревания и поддержание их в течение всей взрослой жизни контролируется только тестостероном. К этим признакам относятся развитие наружных мужских половых органов и придаточных желез репродуктивной системы, усиленное развитие мускулатуры, рост гортани и соответственно более низкий голос, рост и распределение волосяного покрова, особенности поведения, связанные с половой жизнью и родительскими заботами.
20.3.2. Женская половая система
Роль женщины в процессе размножения гораздо больше, чем роль мужчины, и она сопряжена с взаимодействиями между гипофизом, яичниками, маткой и плодом. Женская половая система состоит из парных яичников и фаллопиевых труб, матки, влагалища и наружных половых органов (рис. 20.35).

Рис. 20.35. Упрощенная схема строения женской половой системы. Матка и влагалище изображены в разрезе, а наружные половые органы, отверстие мочеиспускательного канала и анальное отверстие — со стороны поверхности, причем половые губы раздвинуты
Яичники прикреплены к стенке брюшной полости с помощью складки брюшины и выполняют две функции: производят женские гаметы и секретируют женские половые гормоны. Яичник имеет миндалевидную форму, состоит из наружного коркового и внутреннего мозгового слоев и заключен в соединительнотканную оболочку, называемую белочной оболочкой. Наружный слой коркового вещества состоит из клеток зачаткового эпителия, из которого образуются гаметы. Корковое вещество образовано развивающимися фолликулами, а мозговое вещество — стромой, содержащей соединительную ткань, кровеносные сосуды и зрелые фолликулы (см. рис. 20.38).
Фаллопиева труба — это мышечная трубка длиной около 12 см, по которой женские гаметы выходят из яичника и попадают в матку. Отверстие фаллопиевой трубы заканчивается расширением, край которого образует бахромку, приближающуюся во время овуляции к яичнику. Просвет фаллопиевой трубы выстлан ресничным эпителием; перемещению женских гамет к матке способствуют перистальтические движения мышечной стенки фаллопиевой трубы.
Матка представляет собой толстостенный мешок длиной примерно 7,5 см и шириной 5 см, состоящий из трех слоев. Наружный слой называется серозной оболочкой. Под нею находится самый толстый средний слой — миометрий; его образуют пучки гладких мышечных клеток, которые во время родов чувствительны к окситоцину. Внутренний слой эндометрий — мягкий и гладкий; он состоит из эпителиальных клеток, простых трубчатых желез и спиральных артериол, снабжающих клетки кровью. Во время беременности полость матки может увеличиться в 500 раз — от 10 см3 до 5000 см3. Нижним входом в матку служит ее шейка, соединяющая матку с влагалищем. Вход во влагалище, наружное отверстие мочеиспускательного канала и клитор прикрыты двумя кожными складками — большими и малыми половыми губами, образующими вульву. Клитор — это небольшое способное к эрекции образование, гомологичное мужскому половому члену. В стенках вульвы находятся бартолиниевы железы, выделяющие при половом возбуждении слизь, которая увлажняет влагалище во время полового акта.
Развитие яйца у человека (оогенез)
В отличие от образования спермиев, которое начинается у мужчин только при половом созревании, образование яйцеклеток у женщин начинается еще до их рождения и завершается для каждой данной яйцеклетки только после ее оплодотворения. Стадии оогенеза показаны на рис. 20.36. Во время развития плода первичные половые клетки многократно делятся путем митоза, образуя много крупных клеток, называемых оогониями. Оогонии снова претерпевают митоз и образуют ооциты первого порядка, которые остаются на стадии профазы почти до самой овуляции. Ооциты первого порядка окружены одним слоем клеток — гранулезной оболочкой — и образуют так называемые примордиальные фолликулы. Плод женского пола непосредственно перед рождением содержит около 2⋅106 этих фолликулов, но лишь примерно 450 из них достигают стадии ооцитов второго порядка и выходят из яичника (овуляция). Перед овуляцией ооцит первого порядка проделывает первое деление мейоза, образуя гаплоидный ооцит второго порядка и первое полярное тельце. Второе деление мейоза доходит до стадии метафазы, но не продолжается дальше до тех пор, пока ооцит не сольется со сперматозоидом. При оплодотворении ооцит второго порядка совершает второе деление мейоза, образуя крупную клетку — яйцо, а также второе полярное тельце. Все полярные тельца представляют собой мелкие клетки; они не играют никакой роли в оогенезе и в конечном счете разрушаются.

Рис. 20.36. Схема процесса оогенеза у человека
Менструальный цикл
У мужчин образование и выделение гамет — процесс непрерывный, начинающийся с наступлением половой зрелости и продолжающийся в течение всей жизни. У женщин же это циклический процесс, повторяющийся примерно через каждые 28 дней и связанный с изменениями в строении и функциях всей репродуктивной системы. Процесс этот называется менструальным циклом, и его можно разделить на четыре стадии (рис. 20.37). В событиях, происходящих во время менструального цикла, участвуют яичники (овариальный цикл) и матка (маточный цикл), и они регулируются гормонами яичников, секреция которых в свою очередь регулируется гипофизарными гонадотропинами.

Рис. 20.37. Главные фазы менструального цикла (цифры в скобках — дни, на которые приходится данная фаза)
Овариальный цикл
У взрослой женщины овариальный цикл (рис. 20.38) начинается с развития нескольких первичных фолликулов (содержащих ооциты первого порядка) под действием фолликулостимулирующего гормона (ФСГ), выделяемого передней долей гипофиза. Из этих фолликулов только один продолжает расти, тогда как остальные разрушаются в результате дегенеративного процесса (атрезия фолликулов). Клетки гранулезной оболочки растущего фолликула пролиферируют, образуя наружный волокнистый слой толщиной в несколько клеток, называемый theca externa, и внутренний слой, богатый кровеносными сосудами, — theca interna. Клетки гранулезной оболочки секретируют фолликулярную жидкость, которая накапливается в полости фолликула (antrum). Лютеинизирующий гормон (ЛГ), выделяемый гипофизом, стимулирует клетки обеих thecae, побуждая их вырабатывать стероиды, главным образом 17β-эстрадиол. Возрастание уровня эстрадиола во время фолликулярной фазы действует на гипофиз по принципу отрицательной обратной связи, вызывая понижение уровней ФСГ в крови (4-й-11-й дни); уровень ЛГ остается неизменным. Уровень эстрогена достигает максимума примерно за три дня до овуляции и действует в это время на гипофиз по принципу положительной обратной связи, стимулируя выделение как ФСГ, так и ЛГ.

Рис. 20.38. Схема яичника женщины (в разрезе). Показаны последовательные стадии (обозначены цифрами) развития граафова пузырька, овуляции, образования желтого тела и его инволюции. На самом деле нельзя наблюдать все эти стадии одновременно
Как полагают, ФСГ необходим для стимуляции роста фолликулов, но дальнейшее развитие фолликулов контролируется главным образом ЛГ. Гранулезные клетки располагаются по периферии, яйцо смещено к одной стороне фолликула, но все еще окружено слоем гранулезных клеток. Зрелый фолликул, называемый граафовым пузырьком, достигает примерно 1 см в диаметре и выступает над поверхностью яичника в виде бугорка (рис. 20.39). Механизм овуляции в точности неизвестен, но предполагается, что в нем участвуют ЛГ, ФСГ и простагл андины.

Рис. 20.39. Микрофотография зрелого граафова пузырька
Во время овуляции ооцит второго порядка отделяется от стенки лопнувшего фолликула, выходит в брюшную полость и попадает в фаллопиеву трубу. Обычно каждый месяц освобождается только один ооцит одним из яичников, так что овуляция происходит поочередно то в одном, то в другом яичнике. Овулировавший ооцит представляет собой клетку, ядро которой находится в метафазе I мейоза; он окружен слоем клеток, называемым zona pellucida, и слоем гранулезных клеток (corona radiata), который создает защиту ооциту вплоть до оплодотворения.
После овуляции уровень ЛГ падает до уровня, характерного для фолликулярной фазы, и под действием другого гонадотропина — пролактина — клетки лопнувшего фолликула изменяются, образуя желтое тело. Желтое тело начинает секретировать еще один женский гормон — прогестерон — и небольшие количества эстрогена. Эти два гормона поддерживают нормальное строение эндометрия, выстилающего матку, и ингибируют выделение ФСГ и ЛГ, воздействуя по принципу отрицательной обратной связи на гипоталамус. Если оплодотворения не произошло, то под действием факторов, которые будут описаны позднее, желтое тело подвергается инво-люции и от него остается лишь небольшой рубец — corpus albicans; это сопровождается снижением уровней прогестерона и эстрогена, в результате чего прекращается ингибирование секреции ФСГ, его уровень повышается и начинается новый цикл развития фолликула. Соотношения между активностью гипофиза и яичников и изменениями в яичниках и в матке показаны на рис. 20.40.

Рис. 20.40. Общая схема взаимоотношений между гонадотропными гормонами гипофиза (А), стероидными гормонами яичников (Б), развитием фолликулов и желтого тела (В) и толщиной эндометрия (Г) на протяжении менструального цикла женщины
20.8. При вскрытии трупа 22-летней женщины оказалось, что яичники имеют неодинаковые размеры. При исследовании яичников в них было обнаружено:
17 000 фолликулов
4 желтых тела
10 рубцов от желтых тел
25 000 фолликулов
5 желтых тел
48 рубцов от желтых тел
Фолликулы были по большей части мелкие, но 219 имели свыше 100 мкм в диаметре. Пять фолликулов содержали по два ооцита каждый и 2% ооцитов содержали по два ядра. Если предположить, что каждый фолликул образует одно желтое тело, то
1.
а) В каком возрасте у этой женщины начались овуляции?
б) Примерно сколько лет у нее могли бы продолжаться овуляции?
в) Сколько пар близнецов могла бы она родить и какие это были бы близнецы?
2. Чем можно объяснить наличие в некоторых ооцитах двух ядер (см. разд. 22.3)?
Маточный цикл
Маточный цикл делится на три фазы, связанные с определенными структурными и функциональными изменениями эндометрия.
1. Менструальная фаза. В этой фазе происходит отторжение эпителиального слоя эндометрия. Перед самой менструацией кровоснабжение этой области уменьшается в результате сужения спиральных артериол в стенке матки, вызываемого падением уров-ня прогестерона в крови после инволюции желтого тела. Недостаточное кровоснабжение приводит к гибели эпителиальных клеток. Затем сужение спиральных артериол сменяется их расширением, и под действием усилившегося притока крови эпителий отторгается и его остатки выводятся вместе с кровью в виде менструальных выделений.
2. Пролиферативная фаза. Эта фаза совпадает с фолликулярной фазой овариального цикла и состоит в быстрой пролиферации клеток эндометрия, приводящей к его утолщению под контролем эстрогена, выделяемого развивающимся фолликулом.
3. Секреторная фаза. Во время этой фазы прогестерон, выделяемый желтым телом, стимулирует секрецию слизи трубчатыми железами; это поддерживает выстилку матки в состоянии, при котором возможна имплантация в нее оплодотворенного яйца.
Что происходит, если оплодотворение не состоялось?
Если в течение суток после овуляции оплодотворения не произойдет, то ооцит второго порядка претерпевает автолиз в фаллопиевой трубе; то же самое происходит со спермиями, оставшимися в половых путях женщины. Желтое тело сохраняется в течение 10-14 дней после овуляции (обычно до 26-го дня цикла), но затем перестает секретировать прогестерон и эстроген из-за недостаточного уровня ЛГ в крови и подвергается автолизу. Как показали недавние исследования, у некоторых видов стенка матки, не содержащая оплодотворенного яйца, по-видимому, выделяет фактор, названный лютеолизином; этот фактор представляет собой простагландин (простагландин F2α). Как полагают, лютеолизин поступает с кровью в яичник, где он вызывает инволюцию желтого тела, разрушая лизосомы в гранулезных клетках последнего, что приводит к их автолизу.
Результаты оплодотворения
Если происходит оплодотворение, то из образовавшейся зиготы развивается бластоциста, которая через 8 дней после овуляции погружается в стенку матки. Наружные клетки бластоцисты, образующие трофобласт, начинают затем секретировать гормон — хорионический гонадотропин, по своей функции сходный с лютеинизирующим гормоном. Эта функция включает предотвращение автолиза желтого тела и побуждение его к секреции больших количеств прогестерона и эстрогенов, которые вызывают усиленный рост эндометрия. Отторжение эпителиальной выстилки эндометрия подавляется, и очередная менструация не наступает, что служит самым ранним признаком беременности. (Хорионический гонадотропин стимулирует также интерстициальные клетки семенников у зародыша мужского пола и заставляет их вырабатывать тестостерон, который индуцирует рост мужских половых органов.) Роль плаценты возрастает примерно на 10-ю неделю беременности, когда она начинает сама секретировать большую часть прогестерона и эстрогена, необходимых для нормального течения беременности. Преждевременное прекращение активности желтого тела (до того как плацента в полной мере проявит свою секреторную способность) — частая причина выкидыша на 10-й-12-й неделях беременности.
Во время беременности хорионический гонадотропин можно обнаружить в моче, и на этом основаны тесты на беременность. В настоящее время применяется тест, основанный на подавлении агглютинации: частицы латекса, покрытые хорионическим гонадотропином, добавляют к смеси мочи с антисывороткой, агглютинирующей этот гормон. Если он присутствует в моче, то он будет реагировать с агглютинирующей антисывороткой, а не с частицами латекса. Отсутствие агглютинации латекса служит показателем беременности; этот тест можно применять уже через 14 дней после несостоявшейся менструации.
20.3.3. Копуляция
Внутреннее оплодотворение составляет важную часть репродуктивного цикла у наземных организмов, и у многих из них, в том числе у человека, оно облегчается благодаря совокупительному органу — мужскому половому члену, который вводится во влагалище и вносит гаметы как можно глубже в половые пути самки. Эрекция полового члена происходит вследствие локального повышения кровяного давления в его эректильной ткани в результате сужения вен и расширения артерий. Эта реакция обусловлена активацией парасимпатической нервной системы при половом возбуждении. В таком состоянии половой член может быть введен во влагалище, где трение, создаваемое ритмическими движениями во время полового акта, производит тактильное раздражение сенсорных клеток в головке полового члена. Это активирует симпатические нейроны, которые вызывают сокращение внутреннего сфинктера мочевого пузыря, гладкой мускулатуры эпидидимиса, семявыносящего протока и придаточных желез (семенного пузырька, простаты и бульбоуретральных желез). В результате всего этого будущие компоненты семенной жидкости выталкиваются в проксимальную часть уретры, где они смешиваются. Возросшее давление в проксимальной части уретры вызывает рефлекторную активность двигательных нейронов, иннервирующих мышцы у основания полового члена. Ритмические волнообразные сокращения этих мышц проталкивают семенную жидкость через дистальную часть уретры, и происходит эякуляция (выбрасывание спермы) — кульминационный момент полового акта. Ощущения, связанные с этим моментом как у мужчины, так и у женщины, объединяются под названием оргазма. Смазку во время полового акта обеспечивают главным образом секреты желез влагалища и вульвы, а частично также прозрачная слизь, выделяемая бульбоуретральными железами мужчины после эрекции. Секреты придаточных мужских желез имеют щелочную реакцию и содержат слизь, фруктозу, витамин С, лимонную кислоту, простагландины и различные ферменты; они повышают рН среды во влагалище, которая обычно бывает кислой, до 6-6,5, что является оптимальным для подвижности спермиев после эякуляции. Количество семенной жидкости, извергаемой при эякуляции, составляет около 3 мл, из которых лишь 10% составляют спермин. Несмотря на такой низкий процент, сперма содержит примерно 108 спермиев на 1 мл.
20.3.4. Оплодотворение
Сперма изливается глубоко во влагалище, вблизи шейки матки. Как показали исследования, сперматозоиды проходят из влагалища в матку и доходят до верхнего конца фаллопиевых труб за пять минут благодаря сокращениям матки и труб. Как полагают, эти сокращения инициируются окситоцином, выделяющимся во время полового акта, и локальным воздействием простагландинов, содержащихся в семенной жидкости, на матку и фаллопиевы трубы. Спермин сохраняют жизнеспособность в женских половых путях в течение 24-72 ч, но остаются высокофертильными лишь 12-24 ч. Сперматозоид способен оплодотворить ооцит только после того, как он проведет в половых путях несколько часов, претерпевая процесс так называемой капацитации; при этом свойства мембраны, покрывающей акросому, изменяются, что делает возможным оплодотворение, которое обычно происходит в верхней части фаллопиевой трубы.
Когда спермий приближается к ооциту (рис. 20.41), его наружная мембрана, покрывающая область акросомы, и мембраны самой акросомы разрываются, и находившиеся в акросоме ферменты — гиалуронидаза и протеаза — "переваривают" окружающие ооцит клеточные слои, в том числе zona pellucida. Эти изменения в головке спермия называют акросомальной реакцией. В результате дальнейших изменений в головке спермия внутренняя мембрана акросомы выворачивается наизнанку, что позволяет спермию проникнуть сквозь zona pellucida и плазматическую мембрану в ооцит; у человека сперматозоид входит в ооцит целиком. После того как в яйцеклетку проник один спермий, расположенные под ее плазматической мембраной кортикальные гранулы разрываются, начиная с места вхождения спермия, и освобождают вещество, под действием которого zona pellucida утолщается и отделяется от плазматической мембраны яйцеклетки. Эта так называемая кортикальная реакция распространяется по всей клетке, в результате чего zona pellucida образует непроницаемую преграду — оболочку оплодотворения, которая препятствует проникновению в ооцит других спермиев, т. е. явлению полиспермии.
Рис. 20.41 А. Спермии человека, скопившиеся вокруг ооцита второго порядка. Микрофотография, полученная с помощью сканирующего электронного микроскопа. Б. Момент оплодотворения у морского ежа — проникновение спермия сквозь мембрану яйца. Темный клин — головка спермия, в которой заключен генетический материал. Серое образование позади головки доставляет энергию, необходимую хвосту. Спермий переварил наружную оболочку яйца, состоящую из гликопротеина, и проник внутрь. Содержащаяся в яйце жидкость вышла наружу, покрыла спермий и теперь втягивает внутрь, чтобы завершилось оплодотворение. Каким-то неизвестным образом проникновение в яйцо одного спермия закрывает доступ в него для других; вероятно, это связано с быстрыми изменениями наружной оболочки яйца
Проникновение сперматозоида служит стимулом для завершения второго деления мейоза, и ооцит второго порядка становится зрелым яйцом. При этом образуется также второе полярное тельце, которое тотчас же дегенерирует, а хвост сперматозоида рассасывается в цитоплазме яйца. Ядра обеих гамег превращаются в пронуклеусы и сближаются. Мембраны пронуклеусов разрушаются, а отцовские и материнские хромосомы прикрепляются к образовавшимся нитям веретена. К этому времени оба гаплоидных набора, содержащих у человека по 23 хромосомы, уже реплицировались, и возникшие в результате 46 пар хроматид выстраиваются по экватору веретена, как в метафазе митоза. Слияние пронуклеусов называется кариогамией. На этой стадии восстанавливается диплоидное число хромосом, и оплодотворенное яйцо получает название зиготы.
Зигота проходит стадии анафазы и телофазы и завершает свое первое митотическое деление. Следующий за этим цитокинез приводит к образованию двух диплоидных дочерних клеток. Важнейшие события, связанные с оплодотворением, суммированы на рис. 20.42.

Рис. 20.42. Упрощенная схема оплодотворения. А. Спермий после процесса капацитации способен проникнуть сквозь zona pellucida ооцита второго порядка и вызвать разрушение кортикальных гранул. Б. В результате кортикальной реакции zona pellucida превратилась в оболочку оплодотворения, которая отходит от плазматической мембраны, так что между ними образуется перивителлиновое пространство. В. При втором делении мейоза образуется второе полярное тельце, а затем формируются женский и мужской пронуклеусы. Г. Ядерные оболочки пронуклеусов разрушаются; материнские и отцовские хромосомы, прикрепленные к нитям веретена, выстраиваются на экваторе яйца. Это метафаза первого митотического деления зиготы
20.3.5. Имплантация
Проходя по фаллопиевым трубам в матку, зигота дробится (т.е. претерпевает ряд ядерных и клеточных делений) и образует комочек из клеток — морулу. Дробление не приводит к увеличению размеров морулы, так как клетки продолжают оставаться внутри zona pellucida. Получающиеся в результате дробления клетки называют бластомерами; в дальнейшем они располагаются по периферии, образуя стенку вокруг центральной полости бластоцеля, наполненного жидкостью из яйцевода. Наружный слой бластомеров, называемый трофобластом, в одном участке дифференцируется, образуя внутреннюю клеточную массу. Эта стадия развития, достигаемая спустя 4-5 дней после овуляции, называется бластоцистой (рис. 20.43).

Рис. 20.43. Упрощенная схема строения бластоцисты человека спустя четыре дня после оплодотворения
Попав в матку, бластоциста в течение двух дней остается в ее полости; за это время zona pellucida исчезает, так что клетки трофобласта могут вступить в контакт с клетками эндометрия. Клетки трофобласта размножаются, получая питательные вещества из эндометрия, и на 6-й-9-й день после овуляции бластоциста погружается в стенку матки, располагаясь между клетками эндометрия. Этот процесс носит название имплантации. Клетки трофобласта дифференцируются на два слоя; из клеток наружного слоя образуются ворсинки трофобласта, которые врастают в эндометрий (рис. 20.44). Участки эндометрия между этими ворсинками образуют сообщающиеся друг с другом полости, называемые лакунами, что придает этой области эндометрия сходство с губкой. Под действием гидролитических ферментов, выделяемых этой многоядерной структурой, артериальные и венозные сосуды эндометрия разрываются и вытекающая из них кровь заполняет лакуны. На ранних стадиях развития бластоцисты обмен питательными веществами, кислородом и отходами метаболизма между клетками бластоцисты и материнской кровью в стенке матки происходит через ворсинки трофобласта. На более поздних стадиях развития эту функцию принимает на себя плацента.

Рис. 20.44. Бластоциста человека вскоре после имплантации в эндометрий (упрощенная схема). Ферменты, вырабатываемые синцитием наружного слоя трофобласта, разрушают кровеносные сосуды эндометрия, в результате чего образуются наполненные кровью лакуны: через которые бластоциста получает питательные вещества и выводит продукты, подлежащие экскреции
20.3.6. Ранние стадии эмбрионального развития
Наружные клетки бластоцисты, из которых состоит трофобласт, растут и развиваются, образуя наружную оболочку, называемую хорионом; хорион играет важную роль в питании развивающегося зародыша и удалении ненужных продуктов обмена. Во внутренней клеточной массе появляются две полости; клетки, выстилающие эти полости, дают начало двум оболочкам — амниону и желточному мешку. Амнион представляет собой тонкую оболочку, которая покрывает зародыша и несет защитные функции; его клетки выделяют амниотическую жидкость, заполняющую амниотическую полость, расположенную между амнионом и зародышем. По мере роста зародыша амнион расширяется, так что он всегда бывает прижат к стенке матки. Амниотическая жидкость поддерживает зародыша и защищает его от механического повреждения. У человеческого эмбриона желточный мешок не играет существенной роли, однако у рептилий и птиц он поглощает запасенные в желтке питательные вещества и переносит их в среднюю кишку развивающегося зародыша.
Клетки, составляющие внутреннюю клеточную массу, и желточный мешок образуют структуру, называемую зародышевым диском, из которой развивается эмбрион. Клетки этого диска на ранней стадии, когда его диаметр не достигает 2 мм, дифференцируются на два слоя — эктодерму (наружный слой) и энтодерму (внутренний). На более поздней стадии образуется мезодерма, и эти три зародышевых листка дают начало всем тканям развивающегося эмбриона, как это описано в разд. 21.8.
На ранних стадиях развития обмен веществами между зародышем и материнским организмом происходит через ворсинки трофобласта, однако довольно скоро ворсинки уже не могут справляться с этими функциями и из задней кишки зародыша развивается четвертая оболочка — аллантоис. Хорион, амнион, желточный мешок и аллантоис называют зародышевыми, или позднее плодными, оболочками (рис. 20.45). Аллантоис растет в наружном направлении, пока не приходит в соприкосновение с хорионом, образуя богатую сосудами структуру — хориоаллантоис, который участвует в формировании плаценты — органа, более эффективно осуществляющего обмен между зародышем и материнским организмом.

Рис. 20.45. Упрощенная схема взаимоотношений между человеческим эмбрионом и зародышевыми оболочками примерно через 5 недель после овуляции. В области хориона и аллантоиса в дальнейшем образуется плацента
Плацента
Плацента — временный орган, имеющийся только у плацентарных млекопитающих; у животных это единственный орган, состоящий из клеток, происходящих от двух разных организмов — матери и плода. В плаценте кровеносные сосуды материнского организма и плода вступают в тесный контакт между собой, что облегчает перенос питательных веществ, кислорода и отходов метаболизма. Плацента представляет собой обособленное дисковидное образование, расположенное в одном из участков стенки матки; по мере своего развития она принимает на себя функции ворсинок трофобласта и по прошествии 12 недель становится главной структурой, обеспечивающей обмен веществами между матерью и плодом.
Часть плаценты, происходящая от плода, состоит из соединительнотканных клеток хориона, которые инвазируют ворсинки трофобласта и образуют более крупные выросты — ворсинки хориона. Внутренние участки этих ворсинок заполняются сложными сетями капилляров, отходящих от двух кровеносных сосудов плода — пупочной артерии и пупочной вены. Эти сосуды образуются в аллантоисе и идут от плода к стенке матки, находясь в пупочном канатике, или пуповине, — плотном тяже длиной около 40 см, покрытом клетками, происходящими из амниона и хориона (рис. 20.46).

Рис. 20.46. Упрощенная схема взаимоотношений между пупочными кровеносными сосудами, капиллярами ворсинок хориона и наполненными кровью лакунами в плаценте человека. Плацента служит связующим звеном между кровеносными системами плода и материнского организма
Материнская часть плаценты состоит из выростов поверхностных слоев эндометрия — децидуальной оболочки. Между ними и ворсинками хориона расположены лакуны, снабжаемые артериальной кровью по артериолам матки и отдающие венозную кровь по венулам в маточную вену. Направление тока крови через лакуны определяется разницей давления в артериальных и венозных сосудах.
Мембраны клеток в стенках ворсинок хориона, омываемых материнской кровью, образуют микроворсинки, что увеличивает их поверхность, через которую происходит обмен веществами путем диффузии и иными способами. В этих мембранах имеются молекулы-переносчики, служащие для поглощения веществ ворсинками путем активного транспорта; клетки содержат множество митохондрий. То, что в клетках ворсинок много мелких пузырьков, позволяет предполагать, что поглощение веществ из материнского организма происходит путем пиноцитоза (см. разд. 7.2.2). Вероятно, необходимы различные механизмы, так как расстояние, отделяющее кровь плода от крови матери, довольно велико (в десять раз больше, чем, например, расстояние, которое приходится преодолевать при газообмене в легких). Через "плацентарный барьер" от матери плоду передаются вода, глюкоза, аминокислоты, простые белки, липиды, неорганические соли, витамины, гормоны, антитела и кислород, а в обратном направлении — вода, мочевина и другие азотсодержащие отходы, гормоны и СО2. Плоду могут передаваться от матери потенциально вредоносные факторы, такие как бактерии, вирусы, токсины и лекарственные препараты, однако их воздействие нейтрализуется передачей в том же направлении некоторых антител, глобулинов, антибиотиков и антитоксинов; благодаря этому рожденный младенец обладает пассивным иммунитетом к некоторым болезням (см. разд. 14.14.5).
Плацентарный барьер не только защищает плод от последствий многих вредных воздействий, которым может подвергнуться мать, но и экранирует его кровеносную систему от высокого кровяного давления в сосудах матери. Он не может, однако, служить иммунологическим барьером, а плод, поскольку он несет отцовские гены, будет вырабатывать антигены, чужеродные для организма матери, который будет вырабатывать против них антитела. Чем обусловлена удивительная способность плода (в иммунологическом отношении, посуществу, гомотрансплантата) на протяжении 40 недель сопротивляться отторжению, неизвестно; предполагается, однако, что она связана с выработкой иммуносупрессивных веществ, циркулирующих в плазме матери.
Для жизни и развития плода необходимо, чтобы он непрерывно получал от материнского организма кислород; это обеспечивается различным сродством к кислороду гемоглобинов плода и матери (см. разд. 14.13.1).
Плацента представляет собой эндокринный орган, секретирующий главным образом хорионический гонадотропин, эстрогены, прогестерон и плацентарный лактоген (соматомаммотропный гормон). Последний стимулирует развитие молочных желез при подготовке к лактации. Все эти гормоны образуются в соединительной ткани хориона.?
Половое развитие зародыша
Генетический пол зародыша определяется половой хромосомой, содержащейся в отцовском сперматозоиде: если это Х-хромосома, то родится девочка, если Y-мальчик. Однако создается впечатление, что человеческий организм исходно имеет тенденцию к развитию в сторону женского пола, в основном из-за присутствия в клетках обоих полов Х-хромосом. На ранних стадиях онтогенеза у зародыша развивается пара недифференцированных зачатков гонад — половых складок — и образуются зачатки элементов как женской, так и мужской репродуктивной системы. В результате все эмбрионы вплоть до шестой недели развития являются потенциально обоеполыми.
В недавних исследованиях был выявлен возможный механизм, с помощью которого половые хромосомы определяют, какая из двух систем будет активирована и достигнет полного развития у данного индивидуума.
Х-хромосома несет ген Tfm (ген тестикулярной феминизации), нормальный аллель которого детерминирует синтез в клетках развивающейся репродуктивной системы белкового рецептора для андрогена. Поскольку зародыш как мужского, так и женского пола несет по крайней мере по одной Х-хромосоме, это вещество имеется у обоих полов.
Y-хромосома несет особый ген, детерминирующий синтез определенного белка — H-Y-антигена, который стимулирует дифференцировку клеток половых складок в семенные канальцы и интерстициальные клетки. Тестостерон, выделяемый в кровеносную систему зародыша, взаимодействует с рецептором андрогенов в клетках-мишенях потенциальной репродуктивной системы. Образующийся в результате комплекс рецептора с тестостероном переходит в клеточное ядро, где он активирует гены, влияющие на развитие тканей. Тестостерон стимулирует развитие только тех тканей, которые дают начало мужской репродуктивной системе, и поэтому из зародыша XY развивается плод мужского пола. Ткани потенциальной женской репродуктивной системы не активируются и не развиваются. У зародыша XX отсутствие тестостерона позволяет репродуктивной системе развиваться в присущем ей направлении — женском.
Таким образом, можно сделать вывод, что плацентарные влияния направляют развитие в сторону женского пола, если механизм, приводимый в действие Y-хромосомой, не направит его в противоположную сторону. Все это схематически представлено на рис. 20.47.

Рис. 20.47. Общая схема событий, с которыми связана дифференцировка половых складок зародыша и зачатков репродуктивной системы в мужские или женские гонады и репродуктивную систему плода
20.3.7. Роды
С начала третьего месяца беременности человеческий зародыш называют плодом; в общей сложности беременность обычно продолжается 40 недель, по истечении которых наступают роды. На 12-й неделе беременности у плода уже сформированы все основные органы, а остальное время занимает рост.
На всем протяжении беременности секретируются все большие количества эстрогена и прогестерона; сначала их источником служит желтое тело, а затем главным образом плацента. В последние три месяца беременности секреция эстрогена усиливается быстрее, чем секреция прогестерона, а перед самыми родами уровень прогестерона снижается, а уровень эстрогена возрастает. Функции этих гормонов в период беременности представлены в табл. 20.4.

Таблица 20.4. Основные функции эстрогена и прогестерона во время беременности
Раньше считали, что сроки наступления родов регулируются гормонами; однако, как показали результаты недавних исследований на ряде млекопитающих, значительную роль, видимо, играет и плод. Это, вероятно, имеет большой адаптивный смысл, обеспечивая рождение плода на такой стадии развития, когда он способен к относительно самостоятельному существованию. Как полагают, начальные этапы родов вызываются какими-то еще не установленными факторами, заставляющими гипоталамус плода стимулировать секрецию адренокортикотропного гормона (АКТГ) гипофизом. Согласно одному из современных представлений, сигналом для начала родов служит "фетальный стресс", который создается в результате иммунологического отторжения зрелого плода материнским организмом. Выделяющийся АКТГ побуждает надпочечники плода секретировать кортикостероиды, которые, проникая через плацентарный барьер в кровоток матери, вызывают снижение синтеза прогестерона и повышение секреции простагландинов. Снижение уровня прогестерона делает возможным секрецию октапептида окситоцина задней долей гипофиза матери и снимает ингибирующее воздействие на сокращение миометрия. В то время как окситоцин вызывает сокращение гладкой мускулатуры матки, простагландины увеличивают силу этих сокращений. Окситоцин выделяется во время родов "волнами" и облегчает изгнание плода из матки. Начало сокращений миометрия — так называемых "родовых схваток" — сопровождается расширением шейки матки, разрывом амниона и хориона, вытеканием амниотической жидкости через шейку матки и стимуляцией рецепторов растяжения в матке и ее шейке. Импульсы от этих рецепторов возбуждают вегетативную нервную систему, что приводит к рефлекторному сокращению матки. Другие импульсы передаются вверх по спинному мозгу и стимулируют гипоталамус, который побуждает заднюю долю гипофиза к секреции окситоцина. Давление проходящей через таз головки плода раздражает шейку и вызывает более сильные сокращения миометрия.

Рис. 20.48. Гормональные и нервные механизмы, возможно связанные с родами. Из всех приведенных на схеме механизмов твердо установлено только воздействие окситоцина на матку. Цифры 1-7 указывают возможную последовательность событий, вызывающих сокращение стенки матки, а* 8, 9а и 96 возможные рефлекторые реакции, участвующие в регуляции этих сокращений
Сокращения матки распространяются сверху вниз, усиливаясь по мере приближения к шейке и проталкивая ребенка книзу. При этом шейка постепенно расширяется, а перерывы между приступами сокращений уменьшаются. Это первая стадия родов, которая завершается, когда шейка матки достигнет того же диаметра, что и головка плода. На второй стадии головка и тельце проходят через влагалище и младенец рождается на свет. Пуповину перевязывают в двух местах и перерезают между этими местами, полностью выводя ребенка из-под прямой физиологической опеки со стороны матери. В течение 10-45 мин после рождения ребенка матка резко сокращается, плацента отделяется от стенки матки и выходит через влагалище. Это третья стадия родов. Кровотечение при этом ограничивается благодаря сокращению гладких мышечных волокон, которые окружают все кровеносные сосуды, снабжающие плаценту. В среднем потеря крови составляет около 350 мл.

Рис. 20.49. А. Врач, принимающий роды, направляет головку, которая целиком вышла из влагалища

Рис. 20.49. Б. Тельце ребенка наполовину вышло наружу; ножки еще находятся внутри

Рис. 20.49. В. Ребенка уже обтерли, и он лежит на груди у матери. Пуповина пока не перерезана, так что ребенок все еще связан с плацентой

Рис. 20.49. Г. Врач перерезает пуповину, и процесс родов завершается
20.3.8. Лактация
Молочные, или грудные, железы состоят из тканей двух типов — железистой ткани и поддерживающей ткани, или стромы. Клетки железистого эпителия выстилают небольшие мешочки, называемые альвеолами и сгруппированные в молочной железе в 15 долек. Альвеолы окружены слоем сократительной миоэпителиальной ткани, участвующей в выведении молока. Молоко выделяется железистыми клетками и проходит по ряду протоков и синусов, в которых оно собирается к отдельным отверстиям в соске.
В период полового созревания молочные железы увеличиваются в результате развития стромы под влиянием эстрогена и прогестерона. Внутренние изменения характера стромы и молочных протоков, а также количества жира в молочных железах, происходящие на протяжении менструального цикла, приводят лишь к небольшому увеличению молочных желез во время лютеальной фазы, однако во время беременности происходит значительное увеличение размеров желез и их функциональной активности.
Увеличение молочных желез во время беременности связано с развитием их секреторных областей под влиянием эстрогена, прогестерона, кортикостероидов, гормона роста, пролактина и плацентарного лактогена. Однако на всем протяжении беременности наличие в крови прогестерона подавляет образование молока (лактогенез). При родах, когда уровень прогестерона снижается, подавление действия пролактина прекращается и он стимулирует альвеолы к секреции молока.
Женское молоко содержит жир, лактозу (молочный сахар) и белки лактальбумин и казеин; все эти вещества легко перевариваются. Молоко синтезируется из метаболитов, содержащихся в крови: лактоза — из глюкозы под действием фермента лактозосинтетазы, белки — из аминокислот, а жиры — из жирных кислот, глицерола и ацетата. Одного материнского молока достаточно, чтобы младенец ежедневно прибавлял в весе 25-30 г.

Рис. 20.50. Упрощенная схема строения женской груди. Видны дольки, где секретируется молоко, а также протоки и синусы, по которым молоко направляется к соску
В промежутках между кормлениями грудью пролактин стимулирует образование молока для следующего кормления. Для непрерывной секреции молока необходим эстроген; однако с помощью искусственно создаваемого избытка эстрогена можно подавлять лактогенез у матерей, не желающих кормить ребенка грудью (в настоящее время для этого вместо эстрогена используют бромокриптин).
Выделение молока из соска связано с простой рефлекторной реакцией — активным выведением молока. Когда младенец сосет грудь, он стимулирует рецепторы соска, которые посылают импульсы через спинной мозг к гипоталамусу; гипоталамус секретирует окситоцин, выделяющийся через заднюю долю гипофиза. Это вызывает сокращение миоэпителиальной ткани, окружающей альвеолы, и молоко через протоки и синусы выталкивается из сосков.
После родов молочные железы вначале выделяют не молоко, а молозиво. Молозиво желтого цвета, содержит клетки альвеол и богато белком (глобулином), но в нем мало жира. Как полагают, с молозивом ребенок получает от матери антитела, в частности YgA (см. разд. 14.14.3).
20.3.9. Изменения в системе кровообращения плода при рождении
В период внутриутробного развития легкие и пищеварительная система плода не функционируют, поскольку газообмен и питание обеспечивает материнский организм через плаценту. Большая часть насыщенной кислородом крови, возвращающейся к плоду по пупочной вене, обходит печень по венозному протоку, из которого по нижней полой вене идет в правое предсердие (рис. 20.51). Некоторое количество крови из пупочной вены проходит и через печень; поэтому в правое предсердие поступает смесь оксигенированной (насыщенной О2) и неоксигенированной крови. Из правого предсердия большая часть крови проходит через овальное отверстие (в перегородке между предсердиями) в левое предсердие. Некоторое количество крови идет из правого предсердия в правый желудочек, а из него — в легочную артерий, но не поступает в легкие. Вместо этого кровь направляется по артериальному (боталлову) протоку прямо в аорту, обходя легкие, легочную вену и всю левую половину сердца (предсердие и желудочек). Кровь из левого предсердия поступает в левый желудочек и в аорту, которая снабжает кровью голову, верхние конечности, туловище, нижние конечности и пупочную артерию. Давление крови в кровеносной системе плода достигает наибольшей величины в легочной артерии, чем и определяется направление тока крови через плод и плаценту.

Рис. 20.51. Изменения в кровеносной системе, происходящие при рождении (упрощенная схема), ПП — правое предсердие; ЛП — левое предсердие; ПЖ — правый желудочек; ЛЖ — левый желудочек
20.9. Опишите главное изменение, которое произошло бы в кровообращении плода, если бы кровяное давление оказалось наивысшим в аорте.
После родов внезапное расправление легких снижает сопротивление току крови через легочные капилляры, и в них направляется больше крови, чем в артериальный проток; это снижает давление в легочных артериях. Одновременно с этим перевязка пуповины препятствует току крови через плаценту; это увеличивает объем крови, протекающей через тело ребенка, и ведет к внезапному повышению кровяного давления в аорте, левом желудочке и левом предсердии. В результате изменения давления маленькие клапаны, прикрывающие овальное отверстие и открывающиеся в сторону левого предсердия, закрываются, препятствуя току крови по сокращенному пути — из правого предсердия в левое. Через несколько месяцев эти клапаны срастаются с перегородкой между предсердиями и овальное отверстие полностью закрывается. Если отверстие не зарастает, то возникает необходимость хирургического вмешательства для устранения этого дефекта.
Повышенное давление в аорте и пониженное в легочной артерии проталкивает кровь назад по артериальному протоку в легочную артерию и далее в легкие, резко повышая их кровоснабжение. Спустя несколько часов мышцы в стенке артериального протока сокращаются под действием возрастающего парциального давления кислорода в крови и отключают этот кровеносный сосуд. Аналогичный механизм, основанный на мышечном сокращении, отключает венозный проток и усиливает ток крови через печень. Непосредственный стимул, вызывающий отключение венозного протока, неизвестен, но он играет важную роль в преобразовании пренатальной (дородовой) системы кровообращения в постнатальную (послеродовую).
20.4. Филогенез полового размножения у позвоночных
Половое размножение встречается у представителей всех типов животных. Лежащие в его основе процессы гаметогенеза, оплодотворения и образования зиготы в принципе одинаковы во всех группах, однако морфологические, физиологические и поведенческие аспекты полового размножения весьма разнообразны. Многие примеры различий, наблюдаемых среди беспозвоночных, были описаны в гл. 4.
У позвоночных тоже существуют огромные различия во многих аспектах полового размножения; некоторые примеры представлены в табл. 20.5. В эволюции позвоночных можно проследить постепенную адаптацию к жизни на суше. Одна из главных трудностей, которую было необходимо преодолеть при переходе от водного образа жизни к наземному, была связана с размножением. Большинство рыб выбрасывает свои гаметы прямо в воду, оплодотворение у них наружное; яйца содержат значительные количества желтка, у многих видов имеются личиночные стадии, а забота о потомстве встречается редко. У амфибий известен ряд примеров адаптации к наземному образу жизни, однако лишь немногие из них связаны с механизмом размножения. Для размножения амфибии должны возвращаться в воду, и ранние стадии их развития также проходят в воде. Однако у многих видов амфибий наблюдаются весьма сложные формы поведения, связанные с заботой о потомстве. Например, у пипы самец распределяет оплодотворенную икру по спине самки, к которой икринки прилипают, попадая в имеющиеся на ней ячейки, где развиваются головастики. Примерно через три недели молодые потомки соскакивают с материнской спины и переходят к самостоятельной жизни.
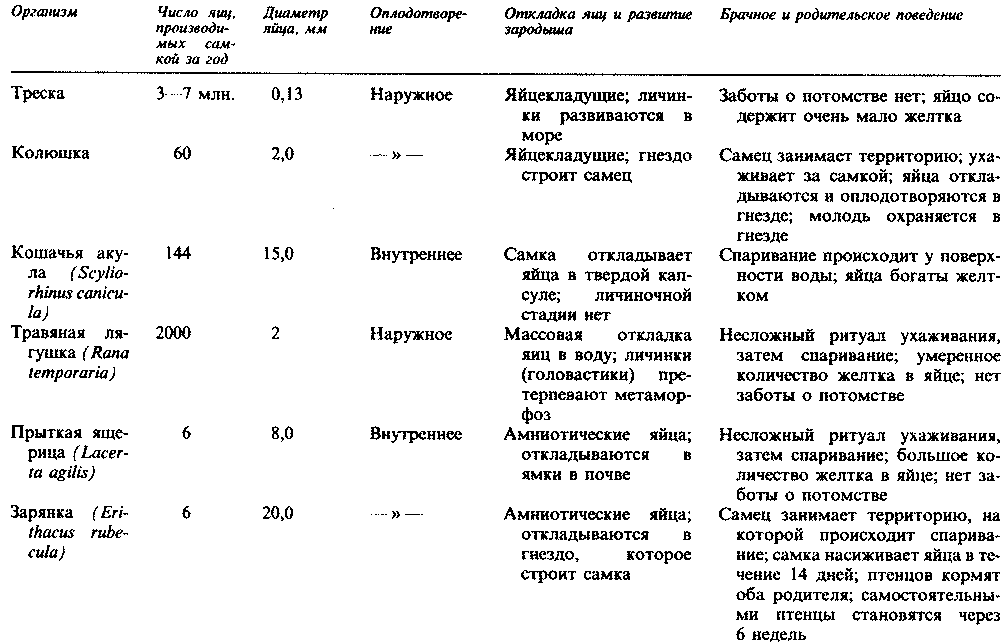
Таблица 20.5. Особенности полового размножения у некоторых позвоночных
Рептилии были первыми позвоночными, разрешившими проблему оплодотворения и развития на суше. Очевидно, что на суше нельзя просто выбрасывать гаметы в окружающую среду, так что первым условием для полного перехода к наземной жизни было введение мужских гамет в тело самки, т. е. внутреннее оплодотворение. Именно такое оплодотворение свойственно рептилиям, и поскольку при этом вероятность слияния гамет возрастает, число их может быть уменьшено. Образующаяся в результате оплодотворения зигота развивается в специализированной структуре — амниотическом (клейдоическом) яйце; в таком яйце эмбрион развивается в полости, наполненной жидкостю. Снаружи яйцо покрыто скорлупой, предохраняющей от механических повреждений. Под скорлупой имеются еще четыре оболочки, окружающие зародыш: желточный мешок, амнион, хорион и аллантоис, происходящие из эктодермы, энтодермы и мезодермы; они защищают зародыш и облегчают осуществление ряда метаболических функций — питания, дыхания, выделения. Желточный мешок закладывается как вырост эмбрионального кишечника; этот мешок обрастает желток, который постепенно поглощается желточными кровеносными сосудами. После того как весь желток использован, желточный мешок спадается и втягивается в кишку. Амнион развивается из клеток, лежащих под зародышем, которые растут вверх и полностью смыкаются над зародышем, образуя амниотическую полость; клетки амниона секретируют амниотическую жидкость, которая заполняет эту полость. Таким образом, эмбрион оказывается в жидкой среде (замена водной среды амфибий), в которой он может развиваться. Амнион имеется у всех рептилий, птиц и млекопитающих, и их называют амниотами. По мере роста зародыша амнион оттесняется кнаружи, пока он не сливается с третьей зародышевой оболочкой — хорионом, которая лежит непосредственно под скорлупой и препятствует чрезмерной потере воды амнионом. Четвертая оболочка — аллантоис — образуется в виде выроста задней кишки; у рептилий и птиц аллантоис быстро разрастается, образуя слой, подстилающий хорион. Он выполняет главным образом функцию "мочевого пузыря", в котором накапливаются отходы метаболизма, и осуществляет газообмен между внешней средой и амниотической жидкостью через пористую скорлупу яйца. Строение яйца амниот схематически показано на рис. 20.52.

Рис. 20.52. Упрощенная схема строения яйца у амниот
Яйца такого типа откладывают также птицы и примитивные яйцекладущие млекопитающие, называемые однопроходными, как, например, утконос (Ornithorhynchus); у высших млекопитающих яйцо утратило скорлупу, но сохранило четыре зародышевые оболочки; у плацентарных млекопитающих две из них, хорион и аллантоис, образуют плаценту (см. разд. 20.3.6).
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги