
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ БИОЛОГИЧЕСКОЙ ФИЗИКИ
с. э. шноль
ФИЗИКО
ХИМИЧЕСКИЕ
ФАКТОРЫ
БИОЛОГИЧЕСКОЙ
ЭВОЛЮЦИИ
ИЗДАТЕЛЬСТВО «НАУКА» МОСКВА 1979
Физико-химические факторы биологической эволюции. Ш н о л ь С. Э.
М., 1979, 263 с.
Рассмотрены физико-химические факторы биологической эволюции, приводящие к возникновению ферментов, аппаратов перемещения в пространстве, систем обеспечения энергией, внутриклеточных структур, клеток, нервной системы. Книга представляет интерес для широкого круга читателей — биологов, физиков, химиков, философов.
Табл. 11, ил. 34, библиогр. 474 назв.
Ответственные редакторы
академик Г. М. ФРАНК, член-корр. АН СССР Г. Р. ИВАНИЦКИЙ
21005—125 2001040000 © Издтельство «Наука*, 1979 г.
055(02)—79
ПРЕДИСЛОВИЕ (ОТВЕТСТВЕННОГО РЕДАКТОРА
В наше время замечательных достижений в биологии поток новых фактов буквально заливает литературу. Быстро изменяются представления о биофизических и биохимических основах явлений жизни. В такой ситуации понятно стремление оглядеться, попытаться представить себе, в какой мере классические основы биологии сочетаются с новыми данными.
Такую попытку и представляет собой книга С. Э. Шноля. Автор исследует в ней возможность дедуктивного метода в современной биологии. С этой целью он прослеживает возможный путь эволюционного становления основных биофизических, биохимических и физиологических свойств живых организмов, а затем кратко рассматривает данные о тех же явлениях, полученные в результате достижений современной науки. Такой подход, интересный сам по себе, позволяет обратить внимание читателя на некоторые узловые вопросы биофизики и биохимии, еще ожидающие своего решения.
Конечно же, не во всех своих разделах анализ автора достаточно последователен и совершенен. Однако основная ценность этой книги — опыт единого в сущности подхода к явлениям жизни на всех этапах биологической эволюции. Автор справедливо подчеркивает, что единственной общебиологической теорией является дарвинизм. Его попытка соединения дарвинизма с современными достижениями биофизики, биохимии, молекулярной биологии в значительной мере есть следствие появления более 50 лет назад классической работы А. И. Опарина «О происхождении жизни».
Книга С. Э. Шноля выходит в свет в период повышенного интереса к проблемам биологической эволюции. Широкую известность, в частности, получила недавняя работа М. Эйгена (1973) «Саморегуляция материи и эволюция биологических макромолекул». Следует отметить, что обе эти работы удачно дополняют друг друга.
М. Эйген сэсредоточил свое внимание в основном на выведении уравнений, связывающих скорость размножения биологических макромолекул данного вида со скоростью и качеством их репликации и гибели.
С. Э. Шноль использует введенное им понятие «кинетическое совершенство» для анализа не только самых начальных, но и последующих этапов эволюции. Примененный им метод («принцип предельного совершенства на каждом этапе эволюционного пути») аналогичен популярному в физике и математике методу предельного перехода.
Следует отметить также, что рассмотрение даже самых серьезных вопросов С. Э. Шноль проводит живо, хорошим литературным языком, не опасаясь местами патетики и даже юмора. Это — хорошо, поскольку показывает до некоторой степени читателк> живую атмосферу научного поиска.
В целом можно рекомендовать эту книгу читателю, интересующемуся проблемами общей биологии, биохимии и биофизики, во всяком случае как источник вопросов, побуждающих думать над их возможными решениями
22 ноября 1973 г.
{Академик Г. М. Франк
Высшим долгом физиков является поиск тех общих элементарных законов, из которых путем чистой дедукции можно получить картину мира.
А. Эйнштейн. Физика и реальность.
Мы унаследовали от наших предков острое стремление к цельному, всеобъемлющему знанию. Само название высших институтов познания— университеты — напоминает нам, что с давних пор и на протяжении многих столетий универсальный характер знаний — единственное, к чему может быть полное доверие...
Э. Шредингер. Что такое жизнь?
ПРЕДИСЛОВИЕ АВТОРА
Широко распространено убеждение в первенствующей роли биологии среди других наук во второй половине XX в. Мысль эта наиболее четко высказывалась физиками (Жолио Кюри, Бернал). Многие выдающиеся физики стали в той или иной форме заниматься биологией и многие внесли в биологию значительный непосредственный вклад (Гамов, Крик, Полинг, Сциллард, Дельбрюк и др.). В высказываниях физиков и в устремлении физиков в биологию проявилась связь наук — то, что наука о природе едина и лишь в человеческом сознании, в силу «невозможности объять необъятное», а также в результате стремления к последовательности и к систематизации знаний она разделена на математику, физику, биологию и др.
Однако дело не только в этом. Именно в физике в начале нашего века произошла общеизвестная революция — были созданы теория относительности и «вантовая механика.
Бурный расцвет новой физики, когда принципиально важные открытия следовали одно за другим с драматической быстротой и приводили к важнейшим теоретическим и практическим результатам, документирован и описан в ряде книг.
Естественно, что именно физики, только недавно пережившие революцию в своей науке, знающие «как это делается», обратились к биологии, где все, как казалось, предвещало аналогичные события.
Их ожидания оправдались. В биологии в самом деле произошла революция, и она действительно явилась результатом прогресса физико-химического направления в биологических ис^ следованиях.
Принято датировать начало революции в биологии 1953 г., когда была опубликована статья Уотсона и Крика о строении ДНК- За прошедшие 20—25 лет преобразились наши знания омолекулярных механизмах наследственности, изменчивости, превращениях энергии в клетке, о механизме биосинтеза основных компонентов протоплазмы — белков, нуклеиновых кислот, липидов, полисахаридов.
Возникла новая наука — молекулярная биология. Но революция в биологии отнюдь не началась статьей Уотсона и Крика. Это было лишь закономерное продолжение открытий Дарвина, Менделя, Моргана, лишь закономерное развитие действительно фундаментальной и революционной идеи XX в. — идеи матричного механизма репликации молекул биополимеров, принадлежащей Н. К- Кольцову. Устремление физиков и физики в биологию действительно оказало большое влияние на темп революционных событий в биологии. Дело не только в проникновении в биологию методов физического и химического исследований. Физика влияет на биологию своим примером, стилем работы, методологией мышления. Именно в физике особо плодотворной оказалась теория. Теоретическая физика замечательна своим ярко выраженным дедуктивным характером, тем, что она является не столько обобщением фактов, сколько развитием исходных общих положений. Триумфы современной физики — это экспериментальное подтверждение предсказаний теории. Получение основных положений какой-либо науки из общих соображений — необходимая завершающая стадия развития мысли в данном направлении. И здесь теоретическая биология сильно отстает от теоретической физики.
Фундаментом современной теоретической биологии служит дарвинизм. Однако после упомянутых физико-химических, «молекулярных» потрясений, после создания «молекулярной биологии» стала ясной недостаточная развитость теоретической биологии. Необходима такая теория биологии, в которой соединены классический эволюционный и новый молекулярный подходы. Возможно по аналогии с физикой и разделение теоретической биологии на макро- и микротеории, однако истинная теория должна быть единой и всеобъемлющей.
Эта предлагаемая вниманию читателя книга — результат убеждения в том, что время для теоретического соединения молекулярной биологии и дарвинизма наступило. Необходимо пытаться получить основные закономерности биологии из общих соображений, построить теорию, позволяющую предсказывать как динамику и траектории эволюционного процесса, так и молекулярные механизмы, функционирующие в биологических системах. Весьма заманчиво получить такие предсказания, а потом посредством эксперимента или обобщения ранее найденных данных сопоставить предсказанное с действительностью. Так биология приблизится к физике в самом главном—в методе научного познания.
Пытаясь рассмотреть биологические закономерности с единых позиций, я прослеживаю далее физические и физико-химическиеусловия начала биологической эволюции и основные критерии естественного отбора в системах полимерных молекул, для которых характерно матричное воспроизведение. При таком исследовании критериями отбора служат физические и физико-химические свойства макромолекул и их комплексов.
Более того, весь процесс биологической эволюции от первичных матричных молекул до высоко совершенных животных и растений можно рассматривать как последовательное, поочередное действие различных физических и физико-химических критериев естественного отбора, т. е. последовательное «включение» различных факторов эволюции.
В настоящее время, конечно, нереально, руководствуясь только общими соображениями, проследить, например, посредством ЭВМ, точные траектории эволюционного процесса. Однако это пока и не нужно. Сейчас вполне достаточен термодинамический подход — выявление лишь начальных и конечных состояний в каждом направлении эволюции. Тогда оказываются ненужными детальные вычисления и становится возможной оценка предельных состояний. Так, если ведущим критерием естественного отбора (т. е. фактором эволюции) при становлении биохимических реакций служит их скорость, легко в принципе оценить, когда дальнейшее совершенствование ферментов станет бесполезным из-за медленности диффузионных потоков реагентов и продуктов. Ведущим критерием отбора, фактором эволюции станет интенсивность перемешивания, и предел возможностей биологической эволюции на данном этапе также можно будет оценить по предельно достижимой скорости перемещения организмов в пространстве. Применительно к биологии такой термодинамический подход выглядит как «принцип предельного совершенства» при анализе основных этапов эволюции.
Можно представить себе опасения, возникающие у читателя книги, озаглавленной «Физико-химические факторы биологической эволюции». Не сводит ли автор биологию только к физике и химии? В сущности, это опасение понятно. И чтобы рассеять его, нужно сначала определить, что такое собственно биология? Не служит ли установление физико-химических механизмов биологических явлений доказательством отсутствия биологической специфики? Нет. Не служит. Биологическая специфика состоит в эволюционном способе (в ходе естественного отбора) формирования всех механизмов и свойств биологических систем. Поэтому собственно биологической является именно теория эволюции. Однако сам ход, этапы, траектории эволюционного процесса вполне подлежат изучению с позиций математики, физики, физической химии (см. нашу статью [340]).
Именно учитывая биологическую специфику, эволюционный способ возникновения всех «механизмов» в биологических системах, автор рассматривает физико-химические факторы эволюции, общие закономерности хода эволюционного процесса, матричного воспроизведения, ферментативного* катализа, превращения энергии, образования дискретных организмов, возникновение ионной асимметрии, появления внутриклеточных структур, механизмы морфогенеза и перемещения в пространстве. Из общих соображений выведена необходимость многоклеточности, наличия рецепторов, нервной системы, мозга.
Естественно, что весь широкий круг вопросов нельзя рассмотреть с равной компетентностью. Я остро ощущаю свой дилета- низм в очень многих затронутых проблемах, но тем не менее, не могу преодолеть привлекательности попытки общего взгляда на современную биологию.
Эта книга сложилась в результате многократного чтения соответствующего курса лекций студентам-биофизикам физического факультета МГУ. Обычно лекции проходили на Беломорской биостанции МГУ. Часто на открытом воздухе, вблизи нового лабораторного корпуса, среди заросших лишайниками валунов, около моря, под шум сосен и крики чаек мы обсуждали общие проблемы биологии. Живая реакция слушателей стимулировала работу над текстом лекций и рукописи. Мне хочется с благодарностью отметить огромный труд директора Беломорской биостанции Н. А. Перцова по созданию этого уникального места для педагогической и научной работы.
Работа над текстом рукописи в ее разных вариантах длилась много лет (с 1967 г.). За это время состоялось множество обсуждений, семинаров, докладов, дискуссий. Особенно полезными были обсуждения затронутых в книге вопросов с Н. В. Тимофее- вым-Ресовским, Л. А. Блюменфельдом, А. М. Молчановым, М. Н. Кондрашовой, Б. Я- Виленкиным, В. И. Дещеревским, Н. Г. Есиповой, Ю. Н. Полянским, М. В. Георгадзе, С. В. Багоц- ким, В. Н. Веховым, В. И. Брусковым, В. Н. Морозовым, Ф. И. Атауллахановым, А. П. Сарвазяном.
Я очень благодарен Г. М. Франку, А. И. Опарину, Г. Р. Иваницкому, Н. А. Райской, В. П. Скулачеву, Э. Э. Шнолю, прочитавшим рукопись и сделавшим ряд ценных критических замечаний. Н. Г. Есиповой написана часть текста гл. 3 и 11. Приложение написано совместно с А. С. Кондрашовым.
Наконец, я должен выразить особую благодарность сотрудникам ОНТИ Института биофизики за неоценимую помощь в подготовке рукописи к изданию.
Работа над текстом этой книги в основном была завершена осенью 1973 г. За прошедшее время некоторые предположения получили независимое развитие в работах других исследователей. То, что казалось экстравагантным, стало обычным. Одной из трудных проблем, возникших в связи с долгим изданием этой книги, оказалось составление списка литературы. Автор чувствовал себя в литературном море как моряк в дырявой лодке — новая литература течет в книгу «сквозь все щели». Дополнения списка литературы лишь отчасти решают эту проблему.
Глава 1
ФИЗИКО-ХИМИЧЕСКИЕ МЕХАНИЗМЫ и биологическая специфичность
Дарвинизм — основа теоретической биологии.
Краткий исторический очерк.
Физические и физико-химические критерии биологического прогресса. Термодинамические характеристики и кинетическое совершенство.
Наиболее важным достижением биологии XIX в. является дарвинизм, лежащий в основе теоретической биологии *. Центральная идея биологии XX в. — концепция матричного молекулярного механизма размножения организмов. Важная задача современной теоретической биологии состоит в выяснении связи матричной концепции с дарвинизмом [17, 23, 24, 129, 262, 288, 289].
Формулирование матричного механизма репликации молекул нуклеиновых кислот как молекулярной основы размножения организмов не только не уменьшает значения дарвинизма, но помогает выявить его действительный фундамент, позволяет понять молекулярные механизмы основных биологических явлений, составляющих предмет дарвинизма. В этом смысле современная молекулярная биология ■— составная часть дарвинизма, теоретической биологии.
Правильность сказанного следует из логического развития следствий матричной концепции. В результате матричного воспроизведения соответствующих молекул в определенных условиях происходят такие процессы: 1) конкуренция размножающихся матричных молекул за вещество, энергию, пространство; 2) вариабельность (изменчивость) матричных молекул; 3) воспроизведение в матричных копиях всех вариантов (конвариантноевоспроизведение, редубликация [288]); 4) естественный отбор, т. е. сохранение и размножение лишь более приспособленных; 5) эволюция, т. е. развитие матричных систем в направлении все большего совершенства, все большей итоговой интенсивности преобразования веществ окружающей среды в вещества данного вида (кинетическое совершенство [339]). Возможность такого в принципе независимого от осуществленного Ч. Дарвиным дедуктивного формулирования дарвинизма посредством анализа молекулярных основ биологии представляет собой важнейшее следствие происшедшей в биологии революции.
1 С современным состоянием и проблемами дарвинизма можно ознакомиться, по статьям, книгам [3, 9, 15, 105, 106, 107, 109, 127, 173, 183, 189, 189а, 199, 202, 241, 252, 289, 290, 304, 332, 333, 359].
Одна из основных задач этой книги состоит в иллюстрации «физико-химической обусловленности дарвинизма» ‘.
Установление обусловленности биологической эволюции фи- зико-химическими свойствами матричных молекул, физико-химический анализ основных ее этапов отнюдь не означает сведения биологии к физике или химии.
Биология отличается ют других .наук тем, что ее объекты представляют результат более или менее длительной эволюции. Они являются, в известном смысле, «концентратами прошедшего времени».
Одна из причин излишних споров «о сводимости» состоит в отсутствии достаточно точных определений основных понятий биологической эволюции, жизни, живого организма.
Я думаю, что настало время попытаться определить их, имея в виду сказанное о дарвинизме как основе теоретической биологии, и о синтезе дарвинизма с молекулярной биологией (см. [339, 340]).
Критерием правильности теории является возможность дедуктивных построений, соответствующих действительности. Поэтому основное содержание предлагаемой книги — опыт дедуктивного построения, попытки «выведения» в качестве следствий из общих, по преимуществу физико-химических соображений основных этапов биологической эволюции, основных свойств живых организмов.
Прежде чем приступить к выполнению этих задач, мне кажется необходимым кратко проследить историю становления концепции матричного механизма размножения организмов. Матричный механизм был сформулирован сначала на основании аналогии между размножением организмов и размножением кристаллов. Аналогия роста (размножения) кристаллов и живых организмов казалась первоначально лишь внешней. Она не имела (и не могла иметь в момент своего возникновения) серьезных оснований. Мысль о такой аналогии возникала на протяжении последних столетий неоднократно. Эту мысль в яркой форме высказывал еще Бюффон (середина XVIII в.). В прошлом веке аналогия с кристаллами была развита Геккелем для описания размножения надмолекулярных агрегатов (пластидул), из которых состоит протоплазма. Л. Я- Бляхер так излагает взгляды Геккеля: «...по мысли Геккеля, пластидулы не размножаются делением, а возникают в питательной жидкости подобно тому, как появляются новые кристаллы под влиянием кристалла, помещенного в насыщенный раствор. Деятельностью пластидул Геккель стремился объяснить размножение, наследственность, приспособление и эволюцию организмов...» [31, с. 31].
Аналогичную концепцию развивал в 1890 г. Альтман, полагавший, что протоплазма состоит из кристаллоподобных гранул; Бляхер пишет о его концепции: «...Альтман высказал предположение, что фибриллярные и сетчатые структуры, как и гомогенная по виду протоплазма, тоже построены из мельчайших гранул-биобластов, которые, по его мнению, являются элементарными единицами жизни. Альтман считал, что гранулы подобно клеткам размножаются делением, почему положение Вирхова «каждая клетка — от клетки» он заменил афоризмом «каждая гранула — от гранулы». Касаясь природы биобластов, Альтман сравнил их с кристаллами, которые по его представлению отличаются от неорганических кристаллов характером роста: неорганические кристаллы растут вследствие отложения вещества из маточного раствора на их поверхности..., тогда как биобласты питаются за счет проникновения надлежащих веществ внутрь» [31, с. 33].
В 1913 г. издательство «Образование» начало выпуск серии книг «Новые идеи в биологии». В первом сборнике этой серии, озаглавленном «Что такое жизнь», был помещен перевод статьи Г. Пржибрама «Обзор мнений авторов о значении аналогии между кристаллами и организмом».
Пржибрам пишет: «...Каждого биолога, углубившегося в кристаллографические изыскания, так сильно поражает «самодеятельность» кристаллов, что граница между неорганическим и органическим миром не кажется ему непреодолимой пропастью..».
Из сходства размножения с кристаллизацией следовали и более широкие биологические аналогии. А. И. Опарин подробно рассматривает эту аналогию в своей знаменитой книге, вышедшей в 1924 г. [227]. В связи со способностью организмов к воспроизведению он пишет[1]: «...указанное свойство присуще не только организмам, но всем без исключения телам, обладающим определенным строением... Возьмем кристалл какого-либо вещества... расколем его на две половинки и бросим последние в перенасыщенный раствор того же вещества. И вот оказывается, что брошенные в раствор половинки кристалла сравнительно быстро восполнят недостающие у них грани, углы, ребра».
А. И. Опарин приводит следующую цитату из книги Каруса Штерне, переведенной с 3-го немецкого издания на русский язык в 1909 г.: «...Здесь происходят самые изумительные явления, которые ...дают ключ к пониманию сложнейших явлений того же порядка. Сера, как известно, простое тело, смотря по температуре, при которой оно переходит из жидкого состояния в твердое, может принимать весьма различный вид — октаэдрическую или призматическую форму. Если опустить два таких кристалла на тонких платиновых проволоках в перенасыщенный раствор серы в бензоле, то по соседству с призматическим кристаллом начинаютобразовываться новые призмы, поблизости же от октаэдрической формы — октаэдры; когда обе армии кристаллов сблизятся между собой, то при первом столкновении последняя форма окажется побежденной. Вот пример борьбы за существование в царстве кристаллов»! [349, с. 200].
Таким образом Карус Штерне в конце прошлого — начале нашего века увидел не только аналогию размножения кристаллов и живых организмов — он обратил внимание на явления «борьбы за существование в царстве кристаллов». Это был очень важный шаг. Для перехода от физической химии к биологии необходимо от «борьбы за существование» кристаллов перейти к естественному отбору. Это становится возможным на основании созданной Н. К. Кольцовым концепции матричного воспроизведения биологических макромолекул [144], концепции конвариант- ной редубликации [288, 467, 468].
Г1о-видимому, именно гипотезы Н. К. Кольцова можно считать идейным началом современной молекулярной биологии. Сам Кольцов [144] утверждал, что мысль о матричном воспроизведении содержалась в докладе съезду естествоиспытателей и врачей, прочитанном А. А. Колли в 1893 г. [141, 142]. На докладе Колли присутствовал студент биологического факультета Московского университета Н. К- Кольцов, развивший потом эту мысль и доложивший свои соображения в 1927 г. на 3-м Всесоюзном съезде зоологов, анатомов, гистологов. У меня эта версия вызывает сомнение, поскольку при чтении текста доклада А. А. Колли мне не удалось усмотреть в нем даже намека на принцип матричного воспроизведения больших молекул. Единственное, что могло послужить толчком для Кольцова в докладе Колли — это вывод о малом числе молекул в бактериальной клетке. Отсюда следовало, что большое число признаков определяется малым числом молекул. Но как именно определяется — это додумал Н. К. Кольцов. Я не могу не привести слова, сказанные им в 1927 г. в своей речи на первом торжественном собрании 3-го Всесоюзного съезда зоологов, анатомов, гистологов в Ленинграде 12 декабря 1927 г.:
«...Процесс «ассимиляции», т. е. точного уподобления вновь возникающих из аминокислот белковых молекул тем образцам молекул, которые уже существуют в данном пункте, является одним из самых загадочных жизненных процессов.
Наиболее простой гипотезой для объяснения ассимиляции является, как мне кажется, заключение, что процесс синтеза белковых молекул сводится к кристаллизации вокруг уже существующих белковых молекул или их агрегатов — кристаллитов, являющихся затравками. Аминокислотные ионы прикладываются своими боковыми сродствами к тем пунктам уже существующих молекул, где находятся соответствующие аминокислоты, совершенно так же, как ионы Na+ и С1“, рассеянные в водном растворе, складываются е определенную решетку вокруг кристалликаповаренной соли... Много понадобилось времени, чтобы установить, что организм возникает только от другого организма из яйца Omne vivum ex ovo, Omnis cellula ex cellula, Omnis nucleus ex nucleo.
Теперь мы можем прибавить еще один новый тезис: каждая белковая молекула возникает в природе из белковой молекулы путем кристаллизации вокруг нее находящихся в растворе аминокислот и других белковых обломков: Omnis molecula ex mole- cula. Значит, размножение не есть исключительное свойство живых организмов, но является наиболее вероятным способом возникновения в природе всех сложных векториальных систем...» [144, с. 482].
Идеи Н. К- Кольцова, опубликованные в 20-е и 30-е годы, были развиты и распространены его учениками. Один из них — Н. В. Тимофеев-Ресовский оказал особенно существенное влияние «а мировоззрение тогда еще молодых физиков, обратившихся к теоретической биологии, например, на М. Дельбрюка {59, 83, 378]. В 1944 г. под впечатлением эгих идей Э. Шредингер написал свою знаменитую книгу «Что такое жизнь» [344]. Ученик Дельбрюка биолог Дж. Уотсон и физик Ф. Крик, занявшиеся биологией под влиянием книги Шредингера, на базе экспериментальных данных М. Уилкинса и Р. Франклин совершили одно из самых важных открытий биологии XX в. — установили структуру молекул ДНК (см. [297, 298]).
Я проследил кратко историю становления матричной концепции для того, чтобы, с одной стороны, показать преемственность идей нашего времени с идеями XVIII и XIX вв., а с другой — подчеркнуть давнее существование теоретической биологии.
Столь же глубоко, а быть может и еще глубже в историю науки уходят и корни эволюционной теории. Подчеркиваю я это потому, что в последнее время вновь стало «признаком хорошего тона» говорить о теоретической биологии в будущем времени как о дисциплине, которую еще лишь предстоит создать. Доля истины здесь, конечно, есть. Теоретическую биологию, как и любую другую теоретическую науку — теоретическую физику например, — отнюдь нельзя считать завершенной.
Однако она существует уже много лет и сейчас не стоит давать названия «На пути к теоретической биологии» книгам, посвященным развитию теоретической биологии (см., например, 1193,194,296]).
Необходимой и достаточной основой теоретической биологии является дарвинизм. Современная молекулярная биология позволяет дать физико-химическую интерпретацию дарвинизму.
Здесь необходимо напомнить, что долгое время совершались (и продолжаются ныне) попытки построить теоретическую биологию не на эволюционной основе, а посредством нахождения особых физических -или химических свойств живой материи. Фи-зико-химическая интерпретация биологических явлений, биохимический и биофизический анализы биологических объектов не имеют, ничего общего с таким неэволюционным подходом к теоретической биологии.
Целеустремленный поиск физического и (или) химического своеобразия биологических объектов был начат в XIX в. Когда прогресс физики и химии определил направление поиска «сущности жизни» в особых химических и физических свойствах веществ, составляющих основу «живой материи». Возникло убеждение, что жизнь — проявление особых свойств особого вещества.
После Лавуазье под влиянием его работ, показавших аналогию процессов брожения, дыхания и горения, распространилось мнение, что жизнь представляет собой особый химический процесс. Сам Лавуазье утверждал, что «жизнь — это химическая функция». По мнению Гегеля, «жизнь есть увековеченный химический процесс» (цит. по [352]). Такое особое, «наделенное жизнью» вещество получило многозначительное название «первичное», «протеин» (отнюдь не соответствующее современному понятию «белок»). Замечательным завершением этого этапа развития теоретической биологии стали широко известные формулировки Ф. Энгельса: «Жизнь — способ существования белковых тел...» [355, 356, с. 77].
Однако отождествление понятий «протеины» и «белковые тела», «протеины» и «белки» не соответствует смыслу определений Ф. Энгельса, что он и сам неоднократно отмечал. В конце XIX в. биохимики выделили из протоплазмы, не отличимой ранее от протеина, ряд существенных фракций. Одна из главных фракций протоплазмы и была названа собственно белком. Тогда же были начаты фундаментальные исследования Э. Фишера, показавшего, что белки — полипептиды, т. е. линейные полимеры, состоящие из аминокислотных остатков, соединенных пептидными
е,ВЯЗЯМИ.
И тут отчасти вследствие семантических неточностей, отчасти из-за стремления возможно быстрее понять сущность жизни, на белки была «возложена» общебиологическая «нагрузка» протеинов. Возникли гипотезы об обусловленности жизни особым составом, состоянием, свойствами молекул белка. Так возникло представление о «живом белке». Поскольку обмен веществ представлялся главным признаком жизни, были предприняты попытки найти причину обмена веществ в особых свойствах молекул белков. Предполагалось, в частности, особое неустойчивое, неравновесное состояние молекул живого белка, допускалось, что спонтанный распад этих молекул служит причиной обмена веществ — обновления молекулярного состава белковых макромолекул *.
Более чем столетний период развития науки в этом направлении завершился появлением книги Э. С. Бауэра «Теоретическая биология» [13]. Бауэр исходил из возможности определения всех основных свойств живых систем посредством анализа физических свойств молекул живой материи.
Согласно Бауэру, фундаментальное отличие живой материи от неживой характеризуется «принципом устойчивого равновесия». Этот принцип гласит: «Все и только живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянно работу против равновесия, требуемого законами физики и химии при существующих внешних условиях». Затем Бауэр в качестве следствий из этого принципа «выводит» основные проявления жизни — обмен веществ, рост, размножение. Бауэр, по-видимому, был неправ, постулируя, что устойчивое неравновесие возникает вследствие особого «напряженного» состояния белковых молекул. «Деформированное» состояние белковых молекул не является основным их свойством. Однако принцип Бауэра верен в эволюционном смысле, а именно, в смысле постоянно возрастающей в ходе эволюции «невероятности», а следовательно, и термодинамической неравновесности биологических макромолекул, «невероятности», проявляющейся в уникальности чередования мономеров в полимерных цепях белков и нуклеиновых кислот.
Сам опыт построения Бауэром теоретической биологии, его логика, конструкция созданной им теории, представляют большую ценность и в наше время. Однако как уже отмечено, его теоретическая биология основана на предположении особых физических свойств живой материи, а именно, особых термодинамических характеристик белков. Бауэр не объясняет эти свойства исходя из каких-либо общих соображений, а «формулирует» их посредством анализа отличий живых организмов от неживых объектов. В этом смысле его подход — неэволюционный. Он признает фундаментальное значение дарвинизма, но не применяет эволюционного подхода для построения теоретической биологии. В этом главный, на мой взгляд, недостаток концепции Бауэра.
Теоретическую биологию можно построить лишь при условии выяснения способа, путей эволюционного становления всех особенностей живых организмов, в том числе становления особенностей химических и физических свойств молекул, составляющих основу живых организмов.
Пути эволюционного становления всех особенностей живых организмов могут быть выявлены посредством применения теории Дарвина с учетом современных достижений генетики, биохимии, биофизики, объединенных в направлении, называемом молекулярной биологией.
Следует отметить, однако, что в последнее время вновь, после Длительного перерыва стали высказываться сомнения в верности дарвинизма (см. [206]). А. А. Любищев классифицировалразличные теории эволюции, полагая при этом дарвинизм лишь одной из нескольких возможных теорий эволюции [185в, г].
С одной стороны, делаются попытки противопоставления теории эволюции в результате естественного отбора случайных мутаций и теории эволюции как закономерного развертывания «внутренних» свойств биологических систем (номогенез Л. С. Берга [21]). Детальное обсуждение этих противопоставлений выходит за пределы задач этой книги (см. [118, 105—107]). Однако противопоставление эволюции в результате естественного отбора и эволюции, как реализации определенных закономерностей совершенно неверно. Закономерность эволюционных траекторий — обязательное следствие, а не противоречие дарвинизму. Поэтому, в частности, неверно рассмотрение закона гомологических рядов Н. И. Вавилова, как противоречие случайной изменчивости и эволюции в результате естественного отбора [21, 206]. Гомологические ряды Н. И. Вавилова лишь подтверждают, в соответствии с теорией Дарвина, закономерный характер эволюции [43, 44, 107]. Гомологичные результаты естественного отбора отнюдь не являются проявлением неслучайного характера изменчивости, а лишь свидетельствуют о действии гомологичных (и даже одинаковых) критериев естественного отбора.
Таким образом, в результате естественного отбора случайных изменений, мутаций, происходит закономерная эволюция. Эволюционные траектории детерминированы, определены физикохимическими и собственно биологическими факторами — это основное содержание настоящей книги. Нет никаких оснований поэтому противопоставлять теорию Дарвина теориям закономерной эволюции. Но при этом речь идет не о таинственных «внутренне присущих системе» законах, а о физических, химических, биологических, доступных рациональному анализу закономерностях.
С другой стороны, высказываются сомнения в какой-либо закономерности эволюции, полагая ее траектории случайным, не зависящим от естественного отбора результатом появления, накопления, комбинации «нейтральных» мутаций [411—414]. «Нейтралистская» концепция возникла при анализе мутационных замен в полипептидных (полинуклеотидных) последовательностях биополимеров [377]. Некоторая часть таких замен не приводит к регистрируемым изменениям функциональных свойств макромолекул. Однако отсюда не следует, что эти нейтральные мутации определяют направления эволюции. Прекрасный анализ «нейтралистской теории эволюции» дан Р. Левонтином. Он пишет: «Никто не настаивает на том, что почти все мутации нейтральны или что эволюция протекает без естественного отбора, главным образом путем случайного закрепления нейтральных мутаций. Оба эти утверждения явно неверны и совершенно чужды духу предлагаемых объяснений. Напротив, мы настаиваем, что многие мутации испытывают действие естественного отбора, но почти всеэти мутации вредны и элиминируются их популяции. Второй распространенный класс представлен группой нейтральных мутаций... Кроме того, рассматриваемая теория допускает существование редких благоприятных мутаций, которые закрепляются естественным отбором, поскольку адаптивная эволюция все же происходит» [173, с. 202].
Из сказанного не следует отрицания физического и химического своеобразия биологических систем. Можно лишь, не претендуя на оригинальность, подчеркнуть, что жизнь, живые существа — результат длительного эволюционного процесса. Живые существа, биологические системы — концентрат многих миллиардов лет эволюции. Биология — наука историческая. Невозможно ни выявить, ни, тем более, понять основные свойства биологических систем вне эволюционного подхода. Только такой подход позволит определить ведущие, характерные признаки живых существ и тем самым позволит определить предмет биологии в целом и каждой ее области в отдельности.
Весь процесс биологической эволюции оказывается возможным подразделить на ряд этапов, отличающихся друг от друга видом, качеством ведущих факторов эволюции. Переход от одного фактора эволюции к качественно другому сопряжен с «кризисом», преодоление которого приводит к резкому ускорению преобразования веществ неживой природы в вещества живых организмов.
Диалектический характер процесса биологической эволюции как процесса, идущего от одного качественного этапа (аромор- фоза) к другому, с преодолением при переходе от этапа к этапу кризисов и противоречий стал ясен из работ А. Н. Северцова [255, 256]. Каждый крупный ароморфоз существенно увеличивает темпы прогрессивной эволюции. В принципе каждый ароморфоз можно количественно охарактеризовать, например, увеличением интенсивности преобразования вещества и энергии у организмов, носителей этого ароморфоза, по сравнению с их предками. Ход процесса эволюции от одного крупного ароморфоза к другому можно рационально описать на основании общих физико-химических соображений. Речь идет в данном случае не о темпах перехода от одного ароморфоза к другому, а лишь о дедуктивном описании возможной последовательности ароморфозов при эволюции в данных условиях.
В некотором смысле такой подход аналогичен термодинамическому, при котором существенны лишь начальные и конечные состояния системы. Аналогию с термодинамикой при анализе процесса биологической эволюции нетрудно продолжить. В самом деле, огромное число микроскопических компонент системы (атомов, молекул), не поддающееся описанию разнообразие траекторий движения и взаимодействий друг с другом этих микро- компонент не препятствие, а условие термодинамического, макроскопического подхода, описания состояний макросистемы. Анало-
тичным образом, огромное число и разнообразие объектов биологической эволюции, чрезвычайная сложность их взаимодействий — не препятствие, а условие описания узловых макроэтапов (ароморфозов) процесса биологической эволюции. На каждом этапе некий существенный в естественном отборе признак, свойство достигает предельного совершенства. Физико-химический и биологический смысл и содержание этого совершенства можно проанализировать без учета времени, необходимого для его достижения (принцип предельного совершенства при анализе эволюционного процесса).
Ясно, однако, что применение принципа предельного совершенства при анализе эволюционного процесса — лишь удобный, а в некоторых случаях и единственно возможный способ первого приближения к действительной картине.
Достаточно полное представление о биологической эволюции может дать рассмотрение не только основных этапов, предельно совершенных по каким-либо факторам, но и темпов эволюции. Далее делается попытка рассмотрения закономерностей, определяющих темпы эволюции, скорость возрастания «биологического прогресса».
Традиционные определения понятий «жизнь, живое существо» были основаны на перечислениях характерных свойств живых организмов. Однако в большинстве случаев удавалось каждое характерное свойство найти в заведомо неживых объектах. Анализ этой ситуации привел к распространенному ныне скептическому отношению к самой возможности таких определений, к убеждению в «бессмысленности» и бесплодности таких попыток {446, 429]. Эту ситуацию четко анализировал Э. Бауэр [13], предложивший, как отмечено выше, в качестве определяющего живое состояние «принцип устойчивого неравновесия».
Затруднения в формулировании необходимых строгих определений снимаются при учете эволюционного происхождения живых организмов: их перечисляемые обычно характерные свойства возникают, формируются в ходе эволюции. В соответствии с этим жизнь — это процесс существования объектов биологической эволюции. Живые существа (организмы) — объекты биологической эволюции [339, 340]. Свойства и характерные черты жизни и жизни существ (организмов) различны на разных стадиях биологической эволюции. Следовательно, единственное, что остается определить, это понятие «биологическая эволюция». Это сделано в следующей главе.
Итогом проведенного далее анализа, по нашему мнению, является доказательство возможности дедуктивного описания важнейших биологических закономерностей и свойств живых организмов посредством рассмотрения процесса эволюции в данных химических и физических условиях. Это значит, что основные этапы биологической эволюции детерминированы физическими и ^химическими факторами окружающей среды.
Глава 2
КИНЕТИЧЕСКИЕ КРИТЕРИИ ЕСТЕСТВЕННОГО ОТБОРА И ФИЗИКО-ХИМИЧЕСКИЕ И БИОЛОГИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Осуществление естественного отбора по признаку кинетического совершенства в системе, способной к полиморфной кристаллизации (матричным синтезам). Принципиальная возможность анализа наиболее вероятных путей биологической эволюции на основе оценки величин кинетического, биологического совершенства и эволюционного потенциала.
Для того чтобы пояснить смысл понятия кинетического совершенства, продолжим аналогию между размножением и борьбой за существование живых организмов и кристаллов.
Представим себе, что некая «жидкость» может с одинаковой вероятностью кристаллизоваться в различных кристаллических модификациях, т. е. имеется система, способная к полиморфной кристаллизации.
В нашем абстрактном случае любая возможная кристаллическая форма имеет одну и ту же вероятность появиться в данных условиях. Отсюда следует, что в этих условиях термодинамическая «выгодность» любой формы одинакова.
В достаточно большом, хотя и ограниченном пространстве, судьба системы, вид, форма кристаллов, заполняющих это пространство, определяется, следовательно, не термодинамическими, а кинетическими особенностями разных кристаллических форм: пространство сначала заполнится формой, которая образуется по случайно возникшей затравке быстрее, чем остальные. Затем по прошествии достаточно долгого времени исследуемое пространство окажется занятым всеми кристаллическими формами в равной доле (поскольку термодинамические характеристики всех форм одинаковы). Представим себе теперь, что более «быстрая», т. е. быстрее заполняющая пространство форма, менее стабильна. И в этом случае такая форма некоторое время может «господствовать» в системе, заполняя большую часть ее объема. Теперь допустим (что вполне реально), что у более быстрой, или в общем случае, у какой-то одной из кристаллических форм расположение молекулярных групп на гранях, сторонах кристалла обусловливает их каталитические свойства, которые позволяют кристаллам данной формы ускорять некую химическую реакцию, сопровождающуюся выделением энергии и способствующую процессу кристаллизации *.
Тогда эта кинетически более совершенная, хотя термодинамически и нестабильная форма за счет свободной энергии, выделяющейся из сопряженно катализируемой реакции, будет все более заметно преобладать над другими формами. Теперь сделаем следующий важный шаг — допустим, что наша система, способная к полиморфной кристаллизации, находится в потоке излучения или каких-то иных внешних воздействий, которые могут стимулировать образование новых вариантов кристаллов, увеличивать разнообразие реализованных форм.
Среди таких измененных, мутантных форм могут оказаться и кинетически более совершенные, т. е, способные к более быстрой кристаллизации и (или) к катализу экзэргонических сопряженных реакций. Ясно, что такие мутантные формы имеют «шансы» заполнить большую часть объема. При этом новом условии — изменчивости — наша система будет развиваться в направлении отбора кинетически все более совершенных форм.
Из изложенных выше соображений следует, что при возможности полиморфной кристаллизации, при наличии разных кинетических (в том числе каталитических) свойств у разных кристаллических форм, при возможности изменчивости кристаллических форм в результате тепловых флуктуаций юли иных причин (мутации), осуществляется естественный (!) отбор форм, способных ко все более быстрому размножению, т. е. к кристаллизации по затравке данного вида.
В результате происходит развитие, смена форм в направлении все большего кинетического совершенства. Эти все более совершенные формы могут (и должны быть) термодинамически все менее вероятными и все более сложными.
Я все время подчеркиваю определяющую роль кинетических факторов в процессе эволюции. Ясно, однако, что любая изолированная система будет следовать термодинамическим законам[2]. И арена эволюции данной системы, ее ареал будет заполнен в конце концов не наиболее совершенными кинетически, а наиболее вероятными, и, следовательно, наиболее термодинамически устойчивыми формами.
Однако в открытой системе при наличии внешнего источника свободной энергии процесс образования и роста матриц может осуществляться сопряженно, т. е. за счет свободной энергии поступающей извне, например, в результате сопряженных химических экзэргонических реакций (см. гл. 7). В таком случае в понятие кинетическое совершенство входит и эффективность сопряжения процесса возникновения и роста матриц (кристаллов) с экзэргоническими реакциями и само «качество» этих реакций. В подобных условиях направление процесса эволюции будут определять не термодинамические, а кинетические свойства кристаллов. Мало того, поскольку отбор будет идти по признаку кинетического совершенства (самого по себе мало вероятного), побеждать в нем будут все менее вероятные формы, будет осуществляться выбор удачных флуктуаций, мутаций. Каждый шаг такой эволюции может быть достаточно вероятным, итог многих последовательных шагов, если забыть об истории, о способе достижения данного этапа, будет казаться совершенно невероятным. Многие исследователи оценивали вероятность возникновения определенной последовательности, например, нуклеотидов в ДНК, и получали величину порядка 10~500. Однако такие внеэволюционные оценки нисколько не противоречат высокой вероятности эволюционного возникновения сложных форм жизни [23, 212, 232].
Действительно, вероятность того, что в данный момент существует какая-либо форма, равна 1. Этого достаточно, чтобы начался естественный отбор (при выполнении указанных выше условий).
Таким образом, естественный отбор начинается с неизбежностью, и вопрос состоит в том, как быстро будет возрастать кинетическое совершенство в эволюционирующей системе. Нужны не оценки вероятности возникновения данной формы (данной последовательности нуклеотидов) молекулы ДНК, а оценки скорости лроцесса эволюции, выяснение того, достаточно ли прошедшего времени для достижения данной «величины» биологического прогресса [264]. Естественный отбор полезных флуктуаций в открытой термодинамической системе с матричным воспроизведением направляет процесс эволюции в сторону, противоположную предписаниям термодинамики закрытых систем. Термодинамика отнюдь не нарушается: процесс эволюции «окупается» сопряженными экзэргоническими процессами. Но направление эволюции определяется кинетическими, а не термодинамическими факторами. Мало того, не просто осуществляется процесс эволюции в направлении все менее термодинамически вероятных форм. Сам лроцесс отбора идет со все большей (до некоторого предела) •скоростью — система не просто удаляется от термодинамического равновесия, а удаляется от него все дальше и с возрастающей скоростью, так как в естественном отборе побеждают более совершенные формы, возникающие быстрее, раньше других. В этом отличие эволюционирующей системы от неэволюционирующей термодинамической системы, где в соответствии с теоремой При- гожина, скорость удаления от равновесия, скорость приращения энтропии минимальна. Естественный отбор, процесс эволюции в Силу давления отбора «заставляет» объекты эволюции с предельно возможной скоростью удаляться от положения термодинамического равновесия, отнюдь не нарушая законов термодинамики. Происходит это за счет использования свободной энергии сопряженных экзэргонических реакций и в результате «ограждения» термодинамически маловероятных структур большими кинетическими (потенциальными) барьерами.
Итак, основной чертой процесса биологической эволюции является определяющая роль, доминирование кинетических, а не термодинамических закономерностей. Это «противотермодинами- ческое» развитие системы в направлении все большего кинетического совершенства осуществляется при возможности использования свободной энергии, освобождающейся при каком-либо экзэргоническом процессе. «Очевидный» механизм эволюционного кинетического совершенствования — случайная или закономерная изменчивость объектов эволюции, сохранение кинетически более совершенных и несохранение менее совершенных вариантов в ходе естественного отбора. Сказанное о естественном отборе в системе с полиморфной кристаллизацией верно для кристаллов любой природы. Однако абсолютная величина интенсивности кристаллизации, предельно возможное кинетическое совершенство зависят от природы кристаллов. Кинетические свойства кристаллов определяются природой элементов кристаллической решетки. Скорость кристаллизации, каталитические свойства поверхности кристаллов зависят от физико-химического разнообразия этих элементов и от величины удельной поверхности кристаллов, поскольку матричный механизм основан в принципе на поверхностном взаимодействии.
Важно подчеркнуть, что кинетические свойства становятся критерием естественного отбора в определенных условиях: при ограниченности «реакционного объема» (возможного ареала), при ограниченности количества вещества, могущего быть превращенным в вещества данного вида, при ограниченности интенсивности потока энергии, поступающей в эволюционирующую систему.
В этих условиях кинетическое совершенство /< определяет скорость преобразования веществ среды в вещества i-ro вида посредством матричных механизмов и задается формулой:
It == rhi = dmjdt = Кти
где nii — масса вещества t-ro вида.
Для характеристики направлений процесса эволюции полезно понятие эволюционного потенциала, определяемого как предельное для данного направления кинетическое совершенство 1тг1.
Эволюция в данном /-м направлении идет до тех пор, пока кинетическое совершенство 1[ не сравняется с /4ах- В таком состоянии исчерпания потенциала система может находиться неопределенное время, пока случайные мутации не «толкнут» ее на новое эволюционное направление. Естественный отбор с высокойскоростью будет усиливать казалось бы ничтожные различия в кинетическом совершенстве разных форм.
Представим себе, сильно упрощая картину, два вида А и В ■(кристаллов, матричных молекул, организмов) скорость размножения которых различна. Пусть они, размножаясь, заполняют предельный объем (ареал), вмещающий ограниченное количество «живого вещества» любого вида. При заполнении этого объема масса вещества разных видов будет пропорциональна соответствующим величинам Представим себе теперь, что ареал заполняется организмами двух видов А и В, затем они живут некоторое время, стареют и умирают, оставляя потомство — зародыши, незначительные по массе, численность которых пропорциональна массе родителей. Жизненный цикл от зародышей до смерти назовем поколением. Легко показать, что соотношение масс организмов видов А и В при 1в=1А-\-г через N поколений будет равно: mA/mB=mAJmBoeNl1. Можно оценить, через сколько поколений N при различных величинах е масса организмов вида А превысит массу организмов вида В в определенное число раз а. Приняв mAJmB = 1, получим:
N = \n(mA/mB) /е.
Эти оценки для наглядности представлены в табл. 1.
Таким образом, 1%-ное различие в величинах е уже через 460 поколений приведет к 100-кратному превышению массы вида А над массой вида В (а=102).
Приведенные оценки не учитывают случайных изменений среды. Флуктуации условий существования могут затруднить или даже сделать невозможным «выход на экспоненту» раз-
таблица 1
ЧИСЛО ПОКОЛЕНИЯ, НЕОБХОДИМОЕ ДЛЯ ПРЕВЫШЕНИЯ МАССЫ ВИДА А МАССОЙ ВИДА В в а РАЗ ПРИ РАЗНЫХ ЗНАЧЕНИЯХ е
е
а
Ю
10*
10»
10*
10»
0,1
23
46
69
92
115
0,01
230
4С0
690
920
1150
0,001
2300
4600
6900
9200
1150
множения особей — обладателей малых селективных преимуществ, когда собственно и начинается естественный отбор. Вопрос об условиях действия естественного отбора во флуктуирующей среде очень сложен. Ему, в частности, посвящены работы Ю. В. Чайковского [319, 321]. Общие закономерности эволюционного процесса с учетом колебания численности популяций, дрейфа генов, флуктуаций, условий существованиярассмотрены в ряде специальных книг ,и статей [57, 66, 80, 82, 105, 106, 108, 122, 127, 173, 189, 202, 203, 213, 235, 241, 243, 264, 289, 290, 308, 316, 332, 333, 326, 327, 375, 387, 351, 412, 468].
Таким образом, приведенные выше численные оценки — лишь иллюстрация высокой эффективности естественного отбора после «выхода на экспоненту», когда небольшие различия в итоговой интенсивности размножения — в кинетическом совершенстве — учитываются естественным отбором.
Является ли кинетическое совершенство действительно универсальным критерием естественного отбора в многосложных процессах биологической эволюции?
Нельзя ли в таком случае «приравнять» термины кинетическое совершенство и биологическое совершенство?
Возможны ли измерения этой величины не только для полиморфной кристаллизации, но и для дальнейших этапов биологической эволюции?
Попробуем ответить на эти вопросы. Действительно, при полиморфной кристаллизации в идеализированных условиях критерием отбора одной из многих равновероятных кристаллических форм является скорость кристаллизации. Чтобы измерить кинетическое совершенство разных кристаллических форм, следует измерить скорость заполнения ими данного объема (ареала) при прочих равных условиях. Процедура ее измерения должна состоять во внесении затравок соответствующих кристаллических форм в способную к кристаллизации жидкость и в определении скорости кристаллизации по данной затравке.
Однако в биологических системах сравниваемые формы могут очень сильно различаться. Можно ли количественно сопоставлять кинетическое совершенство бактерии и слона? Очевидно, нет. Формы, способные строить себя за счет разных исходных веществ, не конкурируют друг с другом за материал. Забравшись в недоступную другим экологическую нишу, некий вид может на более или менее длительное время выйти из-под давления естественного отбора и перерабатывать вещества среды в вещества своего вида значительно медленнее, чем виды, принадлежащие другим направлениям эволюции. Таким образом, Ii служит критерием отбора лишь при прочих равных условиях, при одинаковых условиях существования. Эти прочие равные условия соблюдаются только в момент возникновения новых форм, в самом начале дивергенции, в самом начале вытеснения менее совершенного более совершенным. Дивергентная эволюция приводит к заполнению под давлением естественного отбора всех возможных для данного направления эволюции экологических ниш. В каждой нише рано или поздно окажется вид с предельно возможным в данных условиях кинетическим совершенством. Заполнение всех возможных экологических, ниш А. Н. Северцов [255] назвал идиоадатацией.
Однако в процессе естественного отбора могут возникнуть новые «эволюционные идеи», новые принципы жизнедеятельности, допускающие новую серию эволюционных приспособлений, новую волну заполнения всего разнообразия экологических ниш (в том числе и заполненных ранее). Такие новые принципы приспособления, резкие увеличения эволюционного потенциала Северцов назвал ароморфозами. Старые обитатели различных экологических ниш будут вытесняться новыми (как были вытеснены некогда рептилии млекопитающими, папоротники — цветковыми растениями и т. п.) и их вытеснение будет осуществляться снова по критерию /(— в каждой экологической нише будет разыгрываться своя драма: старый хозяин должен будет сойти со сцены, уступить свое место новому [391].
Следовательно, критерий /< действует при заполнении данной экологической ниши в ходе идиоадаптации, все более совершенного приспособления к жизни в данных условиях, т. е. все большей специализации на базе общего ароморфоза и при вытеснении организмов низшего эволюционного уровня представителями более высокого уровня при появлении нового ароморфоза.
Итак, при прочих равных условиях величина /,• является абсолютной мерой биологического совершенства данного вида. Биологическое совершенство организмов, живущих в разных условиях, можно в принципе сопоставлять лишь с учетом особенностей данных условий существования и внесением соответствующих коррекций в наблюдаемую величину U. Такая коррекция представляет собой в высшей степени трудную задачу даже в случае организмов, занимающих сходные экологические ниши. Поэтому вычисление корректированных, приведенных к нормальным условиям величин позволяющих сравнивать биологическое совершенство бактерий и слонов практически не реально. Это не изменяет того обстоятельства, что величины /,-, приведенные к нормальным условиям, в принципе количественно характеризуют биологическое совершенство данного вида (и вообще, таксономических единиц любого ранга).
Как уже отмечено, Северцову принадлежит открытие общего характера эволюционного процесса. Он писал: «Биологический 'прогресс, т. е. выживание и победа в борьбе за существование, достигается четырьмя различными способами, а именно: 1) путем ароморфозов, 2) путем идиоадаптаций, 3) путем ценогенезов и 4) путем общей дегенерации» [255, с. 100]. Эти способы Северцов определял следующим образом: морфофизиологический прогресс, или ароморфозы — приспособительные изменения, при которых общая энергия жизнедеятельности взрослых потомков повышается; идиоадаптации — приспособительные изменения, при которых энергия жизнедеятельности взрослых потомков не повышается, но и не понижается; ценогенезы — приспособительные изменения зародышей (и личинок), при которых общая энергия жизнедеятельности и строение взрослых потомков не
РИС. 1. Зависимость морфофизио- логического прогресса, общей энергии жизнедеятельности, от времени в ходе эволюции (по А. Н. Се- верцову [256])

Морфофизиологический прогресс
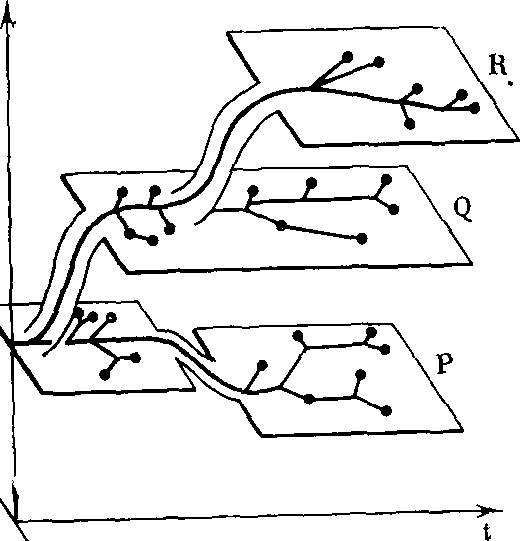
Биологический прогресс может сопровождаться морфофизиологическим ре* грессом (спуск на плоскость Р) ил» морфофизиологическим прогрессом, аро- морфозом (подъем на плоскости Q и- R). В каждой плоскости происходят приспособления к различным условиям, существования (ось со)— идиоадаптации без изменения общей энергии жизнедеятельности; t — время
РИС. 2. Видоизмененный график Северцова, иллюстрирующий возрастание величины кинетического (биологического) совершенства /> в ходе эволюции
Резкие подъемы, ароморфозы, сменяются приспособлениями к разным условиям существования (ось (о) — идио- адаптациями, в ходе которых также наблюдается некоторый рост /,
РИС. 3. Возрастание кинетического- (биологического) совершенства А организмов, приспособленных к разным условиям существования (ось <о) в ходе эволюции
Жирные горизонтальные отрезки — уровни исходных ароморфозов (Р, Q, R), тонкие горизонтальные — уровни / при идиоадаптациях. Вертикальные линии —- переходы от одного ароморфоза к другому. Существование видов с более высоким I исключает существование в тех же условиях (экологических нишах) менее совершенных видов
РИС. 4. Возрастание кинетического совершенства в ходе эволюции при нормировке всех условий существования

изменяются, но возрастает число потомков; общая дегенерация — приспособительные изменения взрослых потомков, при которых общая энергия жизнедеятельности понижается [255, с. 60].
Ход эволюционного процесса Северцов иллюстрировал ставшим широко известным рисунком (рис. 1). В горизонтальных плоскостях происходят идиоадаптационные приспособления к различным условиям существования. Подъемы с одной плоскости на другую представляют ароморфозы, спуски — дегенерацию. Недостатком приведенной схемы является отсутствие строго определенного смысла понятия «морфофизиологический прогресс», «общая энергия жизнедеятельности».
При характеристике хода эволюционного процесса величиной кинетического (биологического) совершенства h схема Север- цова предстает в измененном виде (рис. 2). Эволюция неуклонно идет в направлении возрастания /Биологический регресс (в отличие от морфофизиологического) невозможен. Общая дегенерация (например, у паразитов) становится в этих координатах одним из способов идиоадаптации.
Если для удобства сравнения абсолютных величин кинетического совершенства в одинаковых условиях пренебречь временем, необходимым для достижения данной величины /(, то получится график (рис. 3). При помощи этого рисунка можно сравнивать кинетическое совершенство организмов, последовательно занимавших одинаковые экологические ниши в процессе эволюции. На рис. 3 видно, что новые ароморфозы сопровождаются резким увеличением кинетического совершенства. После каждого ароморфоза происходят относительно небольшие увеличения /,• в ходе идиоадаптаций (сверхтонкая структура спектра /().
Наконец, при нормировке, приведении всех разнообразных условий существования к нормальным, мы получим график (рис. 4). Это безусловно предельное упрощение.
Ясно сознавая сложность такой нормировки, я вынужден буду анализировать далее основные этапы биологической эволюции, имея в виду именно такую предельно идеализированную схему эволюционного процесса.
Сейчас, конечно, нереально пытаться проследить из общих физико-химических соображений путь и темпы биологической эволюции. Однако анализ узловых этапов, основных ароморфо- зов, представляется мне осуществимым.
Появление ароморфоза означает введение в число механизмов кинетического совершенствования очередного нового принципа. Всякий новый ароморфоз становится необходимым по исчерпании возможностей, т. е. эволюционного потенциала, предыдущего.
Эффективность ароморфозов можно в принципе оценить из общих соображений,— посредством вычисления эволюционного потенциала того или иного физико-химического фактора биологической эволюции. К таким факторам относится весь ряд физико-химических механизмов, определяющих кинетическое совершенство. Так, предельная скорость синтеза лимитируется каталитической активностью некоторых компонент системы, скоростью притока и оттока веществ, эффективностью использования свободной энергии в сопряженных экзэргонических процессах, температурой среды и пр., скорость распада матричных молекул зависит от интенсивности внешних излучений,, температуры среды, наличия специальных замедляющих распад приспособлений и т. д.
Все перечисленные выше физико-химические механизмы можно проанализировать, исходя из общих соображений. Следовательно, можно определить (вычислить) и величину эволюционного потенциала каждого очередного этапа, ароморфоза в биологической эволюции.
Я пока не рискую, однако, проводить количественные оценки. В последующих главах я надеюсь показать возможность качественной оценки эволюционного потенциала различных очередных узловых этапов эволюционного прогресса, а также строгую детерминированность самой последовательности основных ароморфозов в эволюции.
Прослеживая динамику событий в абстрактной системе с полиморфной кристаллизацией, способной к конвариантному воспроизведению, мы пришли к выводу, что в ней происходит естественный отбор по признаку кинетического совершенства. В результате этого некая часть системы, та, в которой воплощаются результаты естественного отбора, развивается, становится все более совершенной (и, следовательно, сложной).
Процесс такого развития мы будем называть эволюцией. Эволюцию, идущую в направлении все большего кинетического совершенства, мы будем называть далее биологической эволюцией.
В общем случае не существенна физико-химическая природа веществ, способных к конвариантной редубликации, приводящей к биологической эволюции. Однако среди многих в принципе возможных систем конвариантной (матричной) редубликацииесть лишь одна в принципе предельно совершенная, обладающая максимальным эволюционным потенциалом. По-видимому, такой системой являются нуклеиновые кислоты и белки. Но в данном контексте это не важно. Независимо от конкретной физико-химической природы конвариантно воспроизводящихся матричных молекул, процесс увеличения их кинетического совершенства в результате естественного отбора есть биологическая эволюция.
УВЕЛИЧЕНИЕ БИОЛОГИЧЕСКОГО (КИНЕТИЧЕСКОГО) СОВЕРШЕНСТВА, НАПРАВЛЕНИЕ И СКОРОСТЬ ЭВОЛЮЦИОННОГО ПРОЦЕССА. ПРИНЦИП ДОМИНАНТЫ
Итак, в конкуренции за вещество, энергию и пространство из нескольких «мутантов» побеждает тот, у которого итоговая скорость заполнения возможного ареала выше. Этот, в сущности, кинетический критерий отбора /,• имеет размерность г-сек-1. В самом деле существенна не численность, а количество вещества, преобразуемого в единицу времени в вещества данного варианта (мутанта, вида).
Приведенная выше (стр. 22) зависимость тi от К и т,- основана на кинетике автокатализа, естественной для систем с матричным (типа кристаллизации) механизмом размножения. Величина /< определяется величиной К, являющейся функцией многих переменных величин. В простейшем случае /С,= ЯКц. В частности, К определяется скоростью синтеза мономеров- компонентов матричных макромолекул, скоростью полимеризации уже имеющихся мономеров с образованием макромолекул, эффективностью использования поступающей в систему энергии, устойчивостью образовавшихся макромолекул (организмов) по отношению к неблагоприятным условиям среды и т. д. Соответственно этому величина h является суммой /у, где каждая /13- равна вкладу в итоговую скорость заполнения пространства, т. е. скорости, обусловленной отбором по данному критерию направлению эволюции Kj, т. е.:
Ki = ^ К и и /,■ = VKijtrii. i
Эволюция все время идет в направлении наиболее быстрого возрастания эволюционирующие системы, медленно заполняющие данное «пространство возможностей» будут вытесняться системами, заполняющими это пространство быстрее. Таким образом анализ направлений процесса эволюции сводится к исследованию закономерностей, определяющих скорость возрастания «кинетического» или, что то же самое, «биологического» совершенства U.
Эволюция идет под давлением естественного отбора. Чем больше это давление в данном направлении, тем с большей
вероятностью и скоростью по нему пойдет траектория эволюционного процесса. Замечательно, что повышение скорости движения по данной эволюционной траектории — более быстрое увеличение /; по /-му направлению — означает (в силу конкуренции) одновременное замедление эволюции по другим направлениям. Происходит активное доминирование (аналогично принципу доминанты А. А. Ухтомского применительно к нервной системе [299]) данного направления над другими. В самом деле, носители небольших преимуществ по другим направлениям будут вытесняться в естественном отборе носителями положительных изменений в доминантном направлении, поскольку селективное преимущество оценивается не по вкладу отдельных направлений, а по суммарной величине /4= 2/^.
В то же время давление отбора тем больше, чем ближе по своим свойствам конкурирующие формы. Следовательно, по мере все большего доминирования данной эволюционной траектории, все более узко направленного эволюционного совершенствования скорость возрастания /,■ будет возрастать. Эволюция в данном /-м направлении не означает отсутствия дивергенции. Однако эта дивергенция свойств лишь по данному Кц±Ьц направлению отбора, где ±6* характеризует вариации Кц в данных условиях. По ходу эволюции происходит все более детальная дифферен- цировка этого очередного направления —«разрешающая сила» критерия все время растет. Итак, однажды начавшись, движение по данной эволюционной траектории, т. е. возрастание /« (по К, критерию) идет с возрастающей скоростью, пропорциональной уже достигнутому уровню Л= 2/3- и обратно пропорциональной широте }-го критерия отбора, т. е. /С,±6. Эволюционное совершенствование, т. е. рост А в данном направлении, т. е. за счет роста /з идет до максимально возможного уровня /,• jmax, ограничиваемого физическими, физико-химическими или чисто биологическими причинами. При приближении I{j к IijmdX, среднее приращение АI в результате мутаций становится сравнимым с /шах—Аз, темп эволюционного совершенствования уменьшается и падает до нуля при 1ц=1ц max- Дальнейшее эволюционное совершенствование становится возможным теперь лишь при увеличении /( за счет возрастания по другому направлению — по новому критерию отбора. В силу уже сказанного эволюционное совершенствование по новому направлению сначала идет медленно, а потом «разгоняется». Ход во времени эволюционного совершенствования по каждому отдельному Kj должен иметь вид графика (рис. 5), а при переходе к другому направлению {Kj-О общий вид процесса эволюции во времени также должен иметь вид графика (см. рис. 5).
Вклад в суммарную величину /,• отдельных Itj может быть резко разным, и ход эволюционного возрастания /, представлен на рис. 5.

РИС. 5. Кинетика возрастания Л под давлением естественного оюора в j-u направлении

о—«лаг-фаза», фаза «разгона», фаза «замедления темпа роста по /-му напраилеиню. Длительность лаг-фазы определяется временем, необходимым для установления доминирования /-го направления над другими возможными направлениями эволюции. Фаза разгона— возрастание скорости эволюции по мере сужения ее направления при доминировании. Фаза замедления обусловлена исчерпанием эволюционного потенциала данного направления; б — возрастание 1{ последовательно при эволюции по /-му и (/ + 1)-му направлениям; в — последовательная эволюция по трем направлениям. Показано, что величина приращения I^ при этом может быть различной
Описанная кинетика эволюционного процесса соответствует уравнению:
d/(. ■ _ _ /(Д/,(<)
— 11 — ftii —
it 1 + Б6/(.
В начале движения по данной эволюционной траектории разных мутантов велико. Давление отбора мало. Скорость изменения , мала. По мере продвижения по данной траектории 263ч уменьшается. /f растет (т. е. скорость приращения mt растет). Однако при приближении к тах начинает убывать Д/ (стремящаяся к 0), и эволюция замедляется.
Очередность преимущественного движения эволюционирующей системы в направлении то одного, то другого критерия отбора Kj в принципе может быть установлена из общих соображений [339, 340].
ЭВОЛЮЦИОННО-ТАКСОНОМИЧЕСКИЕ ДИАГРАММЫ
Можно представить себе эволюционное возрастание U как движение по радиусу-вектору в Kj-м направлении (К} задается дугой окружности, измеряемой в радианах, отсчитываемой от направления эволюции в начальный момент времени). Каждому Kj соответствует некоторый диапазон К,+6. Пусть, для определенности, поочередно вступающие в действие критерии отбора (факторы эволюции) располагаются друг за другом на окружности при движении по часовой стрелке. Тогда при данных усилиях процесс эволюции может быть представлен серией «эволюционно-таксономических диаграмм» (рис. 6). Процесс эволюционного совершенствования по данному критерию- состоит в движении по радиусу-вектору в секторе /С,±6. Мерой

РИС. 6. Эволюционно-таксономические диаграммы — изменения во времени Kj — факторы, определяющие направления эволюции (т. е. увеличение 1$, расположены по окружности, по часовой стрелке и измеряются отрезком дуги окружности в радианах. Кинетическое (биологическое) совершенство измеряется суммарной площадью диаграммы; а, б, в, г, д — различные этапы эволюции

РИС. 7. Эволюционно-таксономические диаграммы трех (абстрактных) близких таксонов (видов, родов и т. п.)
эволюционного совершенствования в /-м направлении является площадь соответствующего сектора. Полной мерой биологического (кинетического) совершенства является суммарная площадь всех заполненных секторов (заштрихованных на диаграммах). По завершении эволюции по всем направлениям достигаются максимально возможные в данных условиях величины площади соответствующих секторов. (Эти предельные значения могут быть, как уже отмечалось, резко различными для разных Кц).
В таком состоянии темп эволюции резко замедляется, и естественный отбор становится лишь «стабилизирующим» [332]. В других условиях величины /j, шах будут иными — большими или меньшими, чем в данных. Соответственно иной будет и соответствующая диаграмма. Организмы, соответствующие разным эволюционным диаграммам (разным «экологическим нишам»), будут относиться к оазным таксономическим группам (рис. 7)
РИС. 8. Эволюционная диаграмма при нормировке приращений U по разным Kj
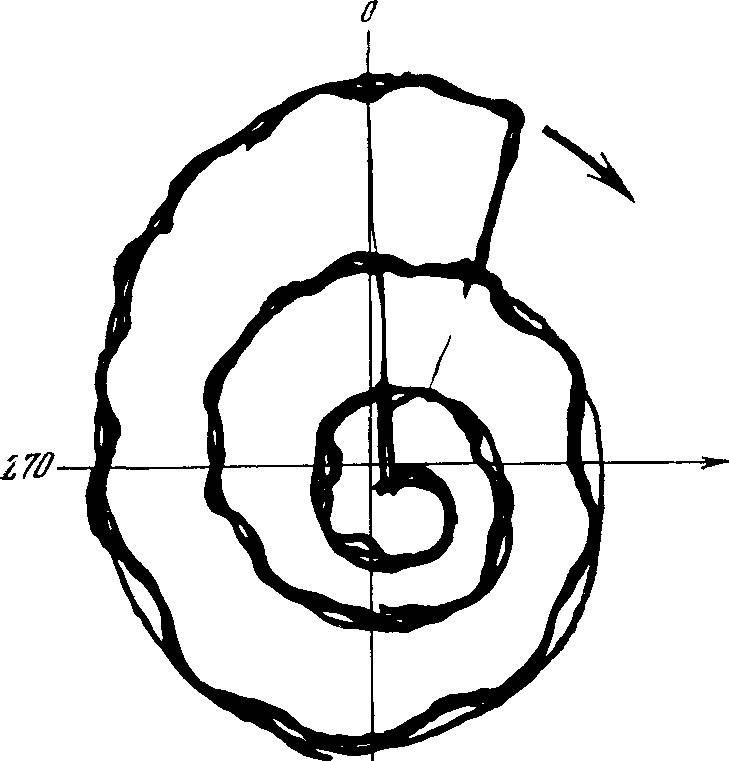
Каждый новый виток спирали осуществляется в результате ароморфоза — возникновения возможности нового возрастания Iij по уже пройденным ранее направлениям. 1( измеряется площадью диаграммы
т
в зависимости от ранга отличия Kj к разным видам, разным родам и т. д.
Анализ темпов эволюции соответственно по формуле (1) и диаграммам приводит, таким образом, к выводу о резко неравномерном темпе эволюционного совершенствования — быстром изменении площадей диаграмм до достижения предельных значений Ij замедлением скорости роста 1{ при переходе от Kj-го к Kj+i-му критерию отбора, и, наконец, неопределенно долгой задержке эволюционирующей системы в состоянии U, тах= 2/3-1тах (см. о неравномерности темпов эволюции работу Н. Н. Иорданского [122]).
Это свидетельствует в пользу реальности устойчивых дискретных таксономических единиц видов, родов [105, 189, 201, 202, 203].
Представленная выше кинетика эволюционного процесса основывалась на предположении, что lit ша* неизменны в данных условиях. Однако вполне возможно изменение /3-, тах и при постоянных внешних условиях в результате достижения некоторой степени совершенства по другим Kj+m-направлениям. С этого момента вновь становится возможным эволюционное совершенствование по ранее пройденным траекториям. Если эти изменения затрагивают все пройденные ранее направления, становится возможным «второй виток» эволюционной спирали»—начинает увеличиваться площадь диаграммы U в тех же, в принципе, внешних условиях (рис. 8). Такие изменения Ij по некоторым К, которые делают возможным возрастание /тах по другим /С,, можно назвать «ароморфозы» в соответствии со смыслом этого понятия, предложенного А. Н. Северцовым. [255, 256]. Таким
33
2 С. Э. Шнольобразом, ароморфоз делает возможным новый виток эволюционной спирали — заполнение (в том числе ранее заполненных) экологических ниш организмами с большей величиной 1тах.
ФИЗИКО-ХИМИЧЕСКИЕ И БИОЛОГИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Критерии, по которым осуществляется естественный отбор, т. е. различные Kj являются, в сущности, факторами эволюции (от facere — делать — лат.). Они определяют направления эволюции. Как мы видели, действие этих факторов происходит последовательно. Очередность действия этих факторов, как отмечено выше, может быть установлена на основе «принципа предельного совершенства» [339, 340].
Начальные этапы биологической эволюции определяются преимущественно физико-химическими факторами. Затем возникают, становятся критериями естественного отбора и собственно биологические факторы.
Биологическая эволюция начинается с началом естественного отбора матричных макромолекул, способных к конвариантному воспроизведению. Критериями отбора вариантов таких макромолекул являются их каталитическая активность, совершенство сопряженных процессов превращения энергии, скорость и экономичность превращения веществ, совершенство кодирования, записи и считывания наследственной информации, совершенство хранения (сохранения) наследственных текстов. Этот путь, по существу, чисто биохимической эволюции ограничен диффузионным барьером— медленностью диффузии метаболитов и ката- болитов. В связи с этим начинают доминировать факторы, направляющие эволюцию по пути ослабления и преодоления диффузионных ограничений. В результате возникают сложные морфологические структуры и аппараты активного перемещения в пространстве. Эволюция, направляемая отбором по критерию все более быстрого и совершенного перемещения в пространстве, приводит к возникновению мышц, нервов, рецепторов, центральной нервной системы. Несмотря на сложность результатов, например, возникновение нервной системы, критериями отбора, приведшими к ним, являются чисто физико-химические факторы эволюции.
Однако уже на этих этапах в действие вступают и собственно биологические критерии естественного отбора. Характерным свойством собственно биологических факторов биологической эволюции является то, что они сами возникают в результате биологической эволюции. К ним относятся прежде всего критерии, определяющие соответствие организмов в биоценозах. Действительно, предметом внимания естественного отбора оказывается и взаимное соответствие разных организмов в биоценозах. Общеизвестным примером эволюции, направляемой
собственно биологическим критерием отбора, является сопряженная эволюция цветков и опыляющих их насекомых, средств нападения хищников и защитных приспособлений жертв, симбиотических водорослей и грибов в лишайниках и т. п.
Мы видели, что скорость эволюционного совершенствования в данном направлении также находится под давлением естественного отбора. Если возможны несколько траекторий, ведущих к данной величине биологического совершенства, выбирается та, которая приводит к конечному этапу быстрее. Следовательно, наряду с факторами, обеспечивающими максимально высокую итоговую скорость конвариантной редубликации, ферментативного катализа, наибольшую эффективность превращения энергии, наибольшее совершенство морфогенеза, перемещения в пространстве, переработки информации — должны существовать •специфические факторы, определяющие (максимально возможную) скорость эволюционного совершенствования на каждом, этапе.
Факторы, определяющие скорость эволюции, могуть быть внешними и внутренними по отношению к эволюционирующей -системе. Внутренние — это те, которые сами возникают в результате эволюции. Это также собственно биологические факторы ■биологической эволюции. Скорость эволюционного совершенствования, как мы видели, зависит от узости интервала Д/С,- ч величины ДU. Биологические факторы в ходе естественного ■отбора по преимуществу влияют на величину Д/, т. е. на величину приращения U за некую единицу времени. А/ зависит от частоты мутаций, скорости размножения, скорости накопления полезных мутаций, эффективности действия естественного •отбора. Рассмотрим эти зависимости подробнее.
Частота мутаций определяется как внешними факторами, в частности, интенсивностью ультрафиолетовой и ионизирующей радиации, так и активностью биохимической репарационной •системы [61]. Ясно, что максимальная скорость эволюции достигается при поддержании частоты мутаций в некотором диапазоне значений, т. е. что для данных условий должна поддерживаться оптимальная частота мутаций.
Скорость увеличения биологического совершенства, т. е. скорость эволюции, возрастает при переходе от бесполого размножения к половому. Действительно, при бесполом размножении скорость возрастания биологического совершенства •определяется скоростью накопления в геноме полезных мутаций. Этот процесс зависит от вероятности мутирования и времени, необходимого естественному отбору для установления полезности или вредности данной мутации. Большинство «признаков», свойств живых организмов формируется в результате сочетания большого числа мутаций. Мутации происходят независимо друг •от друга. Полезной данная мутация может оказаться обычно лишь при условии предварительного осуществления и отбораряда других. Тем самым становится вероятной такая ситуация. В данном множестве родственных организмов имеется весь комплект случайно распределенных между особями потенциально полезных мутаций. Однако особи-носители большинства этих потенциально полезных мутаций гибнут в естественном отборе. Их полезность не проявляется в отсутствии полного комплекта всех предыдущих. Эволюция идет посредством отбора носителей первой полезной мутации и гибели всех прочих, ожидании появления второй полезной мутации у обладателей первой и т. д. На каждом этапе происходит «безрассудное уничтожение ценностей». Еще ярче это выявляется во вполне вероятной ситуации, когда никакая из мутаций в отдельности, без полного их комплекта, не дает преимуществ их обладателя в естественном отборе. Полный комплект данных мутаций очень полезен — сильно увеличивает кинетическое совершенство. В этом случае можно представить себе неопределенно долгую задержку эволюции на одном месте.
Положение резко изменяется при половой рекомбинации геномов. Теперь становится относительно быстро достижимым сочетание мутаций, резко увеличивающих биологическое совершенство обладателей всего комплекта. Правда, половая рекомбинация обеспечивает как сочетание генов разных особей, так и нарушение уже сложившихся полезных сочетаний. В этом случае для «дела эволюции» полезно чередование полового и бесполого размножения. При следующем за половой рекомбинацией бесполом размножении возникает множество копий, пусть еще незавершенного, по полезного набора генов. Испытание в ходе «борьбы за существование» особей, возникших при бесполом размножении, позволяет существенно увеличить концентрацию полезных сочетаний генов (мутаций). Так илн иначе, но и при половой рекомбинации — процессе перемешивания различных геномов—может потребоваться очень большое время, долгий ряд поколений. Все это время носители потенциально полезных мутаций должны сохраняться в системе, быть участниками общего «обменного фонда» генов. Целостность, сохранение этого обменного фонда на протяжении всего времени перебора сочетаний потенциально полезных мутаций—важное условие возможно быстрого возрастания биологического совершенства соответствующих организмов. Наибольшую опасность для сохранности, целостности данного обменного фонда генов представляет его преждевременное разбавление путем половой рекомбинации с представителями других обменных фондов. Поэтому условием ускорения эволюционного совершенствования оказываются изолирующие механизмы, все механизмы, препятствующие размыванию границ данного обменного фонда генов.
Итак, существование живых организмов в виде дискретных видов является условием ускорения эволюционного прогресса.
Пока мы пренебрегали колебаниями, случайными или периодическими долгопериодными изменениями условий существования. Я^сно, что может быть два способа эволюции в переменных условиях существования: 1-й способ — достижение такой скорости генетического приспособления, при которой эволюционные изменения успевают следовать за колебаниями условий существования; 2-й способ — демпфирование — нереагирование на относительно быстрые колебания условий существования, выработка приспособлений, позволяющих жить в соответственно широком диапазоне условий, т. е. такой темп генетических изменений эволюционного совершенствования, который обеспечивает увеличение биологического совершенства, учитывающее лишь постоянную составляющую изменений внешней среды.
му способу соответствует гаплоидный геном, когда каждая мутация сразу же проявляется в фенотипических свойствах ее носителя, сразу же становится объектом естественного отбора.
му способу соответствует полиплоидный (обычно диплоидный) геном, когда «на всякий случай» в каждом геноме имеются не проявляющиеся без должных сочетаний рецессивные гены.
2-й способ эволюции при наличии рецессивных и доминатив- ных алельных генов возможен, как ясно, лишь при половой рекомбинации, когда в гомозиготных организмах проверяется полезность данного гена, а в гетерозиготных — рецессивные гены «ждут своего часа» (см. [65]).
Можно представить себе, что в зависимости от условий существования наибольшая скорость эволюции может обеспечиваться иногда лишь весьма сложной стратегией: сменой гаплоидных и диплоидных поколений, чередованием полового размножения с неполовым, например, партеногенетическим.
Обсуждаемые в этой главе вопросы — сопоставление скорости эволюции при бесполом и половом размножении, образовании дискретных видов, гаплоидности и диплоидности, определенной очередности полового, бесполого, гаплоидного, диплоидного поколений — все это предмет для математического анализа на ЭВМ. Однако до последнего времени сделаны лишь первые шаги в этом направлении. При этом необходимо отметить, что еще[3] в 1930 и 1932 гг. Р. Фишер [387] и Меллер [432, 433] высказали ряд глубоких соображений о зависимости скорости эволюции от половой рекомбинации. Эти соображения близки приведенным нами выше. Однако строгий математический анализ с моделированием и исследованием посредством ЭВМ этой проблемы только начат. В работах Кроу и Кимуры [372, 373], Мейнарда Смита [424] и Фельзенстейна [386] исследуется вопрос о величине ускорения эволюции в результате половой рекомбинации при заведомо упрощенных предположениях, (см. также [450]). Даже при весьма сильных упрощениях Мейнард Смит [424] и Фельзенстейн [386] пришли к выводу о значительном ускорении эволюции при наличии половой рекомбинации. Мейнард Смит оценивал число поколений, необходимое для накопления примерно 10 полезных мутаций (независимых друг от друга) при бесполом и половом размножении. Он получил, что при размере популяции 109 пользе от каждой такой мутации в 0,01 (относительного ускорения размножения) и вероятности возникновения мутаций 10"9, эволюция ускоряется примерно в 6 раз. Сходные, в принципе, оценки получил Фель- зенстейн. Этот автор, кроме того, провел анализ на ЭВМ высказанного Меллером предположения о весьма быстром ускорении эволюции не столько от увеличения скорости накопления полезных мутаций, сколько от эффективного освобождения генома при половой рекомбинации от вредных мутаций. Проведенный анализ, имеющий явно предварительный характер, подтверждает предположение Меллера.
Можно надеяться на быстрый прогресс в этой области. Необходим математический анализ разных способов биологической эволюции, биологического смысла гаплоидности, диплоид- ности, дискретности видов, смены поколений.
Свидетельством реальности этих надежд можно считать работы В. В. Меншуткина и Б. М. Медникова по математическому, посредством ЭВМ, анализу эволюционных закономерностей и, в частности, процесса дивергентного видообразования [408—410].
Об исследованиях М. Эйгена и задачах книги. Во время работы над рукописью книги были опубликованы исследования М. Эйгена, посвященные проблемам самоорганизации материи и эволюции биологических макромолекул [350] *. Они производят большое впечатление ясностью и смелостью постановки и разработки принципиальных вопросов теоретической биологии. Мне представляется, кроме того, весьма интересным, что в ряде пунктов наши позиции оказываются вполне сходными. Такое сходство, по-видимому, имеет особый смысл: отталкиваясь от разных исходных положений, будучи специалистами в разных областях, основываясь на разных ассоциациях и научной предыстории, мы приходим к ряду аналогичных выводов. Поэтому ниже будут кратко рассмотрены некоторые основные аспекты работы Эйгена и отмечены сходство и различия наших взглядов.
Задача книги Эйгена формулируется им так: «Если мы хотим уничтожить разрыв между физикой и биологией, то необходимо разобраться в том, что такое «отбор» на языке точных молекулярных понятий, которые в конечном счете могут описываться квантово-механической теорией. Мы должны вывести дарвиновский принцип из тех свойств материи, которые нам известны» ([350, с. 21]). Рассмотрев необходимые предпосылки для осуществления отбора, Эйген пишет феноменологическое кинеги-
ческое уравнение для изменения численности каждого носителя информации (в нашей терминологии вида):
Xi = (Ft — Ri) Xt + Yi фахи i*i
где х{ — концентрация i-ro носителя информации, — определяет скорость «самоинструктированного» (в нашей терминологии матричного) его образования, R( определяет скорость уменьшения его концентрации (посредством распада, разбавления и т. д.). Последний член (сумма) уравнения характеризует все другие каналы изменения хи которое происходит в результате неточного копирования других матричных последовательностей, близких к i-той, фи определяет скорость образования таких копий.
Это уравнение описывает типичный автокатализ с увеличением численности л:4 при F^Rt. Величины Ft и Rt можно представить в виде:
Ft = kgAjQf, Ri = k0Di + Фиь
где — константы скорости копирования i-й матрицы, Qi—«фактор качества», показывающий, какая доля образовавшихся матриц представляет собой точные копии i-й, k„D — константы скорости уменьшения концентрации i-й матрицы, cpot имеет смысл, аналогичный ф«. С учетом этого основное уравнение Эйгена принимает вид:
Х{ =• kQ — Д] Xi + ^ ф цХ1 — ФоЛ.
Здесь весьма существенно замечание Эйгена, отмечающее, что это уравнение еще не описывает процесса отбора. Для отбора, для настоящей конкуренции за выживание необходимы внешние селекционные ограничения.
При ограничении суммарной численности всех N носителей информации (в нашей терминологии определенный ареал, ограниченные потоки веществ и энергии) получается следующее. Пусть —Д характеризует избыточную продуктивность;
тогда средняя продуктивность будет равна:
_ .V j N
Е = ^ Ekxk / ^ хк• k—i I k=i
Величину —Д Эйген называет селективной цен
ностью, заимствуя этот термин из работ Фишера, Райта, Холдена. После соответствующих преобразований Эйген получает основное уравнение, характеризующее динамику отбора:
Xi = k0 [Wi — Ё] Xi + >] фaxi.
ш
Теперь будет расти численность только тех носителей информации, _селективная ценность Wt которых оказывается выше порога Е. Здесь, вероятно, наиболее важно то обстоятельство, что средняя продуктивность Ё во всей системе непрерывно растет в ходе эволюции до тех нор, пока не будет достигнут ее оптимум, равный максимальной селективной ценности Wm, возможной для данной системы видов. Приближение Е к Wm совершенно аналогично рассматриваемому мною исчерпанию эволюционного потенциала в данном направлении эволюции.
Состояние с Wm на некотором этапе эволюции Эйген рассматривает как селективное равновесие и указывает, что оно имеет метастабильный характер. Это равновесие сохраняется до тех пор, пока не возникнет селективное преимущество с Wm+l>Wm. Как только появляется новая копия такого рода 4i+1, прежнее равновесие разрушается, и система идет к новому (метастабильному) состоянию равновесия.... ([350, с. 52]). Здесь мы видим явную аналогию с появлением очередного ароморфоза.
Далее Эйген анализирует стохастический подход к отбору, подчеркивая ограничения детерминистической теории отбора. По его мнению, элементарный процесс, ведущий к возникновению какого-либо конкретного мутанта, существенно недетерми- нирован, и автокаталитическое усиление ведет к макроскопическому отображению случайных микроскопических событий. Кроме того, он отмечает, что процесс роста численности сам по себе подвержен статистическим флуктуациям и указывает, что определенные стационарные состояния в отличие от истинно равновесных метастабильны и не могут стабилизироваться, и поэтому для поддержания их в течение длительного времени необходима регуляция [350, с. 75]. С учетом отмеченных ограничений Эйген проводит стохастическое рассмотрение процессов отбора.
Эти вопросы и особенно возможную недетерминированность направлений эволюции в результате автокаталитического усиления случайных событий, я рассматриваю в гл. 13, где на основании данных по биологической конвергенции прихожу к другим выводам.
Затем Эйген дает очень интересный анализ возможности эволюционного совершенствования, основанного на комплементарном узнавании только нуклеиновых кислот, и повторяет его для чисто белковых каталитических систем. Он приходит к выводу, что ни нуклеиновые кислоты, ни белки-ферменты не обеспечивают осуществления естественного отбора. Эйген рассматривает также систему, образованную нуклеиновыми кислотами и белками, в которой сочетается комплементарное инструктирование (матричное конвариантное воспроизведение в нашей терминологии) с каталитической связью. Эта система — само- воспроизводящийся гиперцикл — может эволюционировать,т. е. совершенствоваться, путем использования селективных преимуществ. Замечательно, что для гиперциклов с тремя и более членами теоретический анализ предсказывает колебания [350, ,с. 151] что, как отмечено мною, является 'необходимым условием ускорения эволюции.
Далее весьма кратко Эйген отмечает необходимость «ком- партментализации», т. е. образования дискретных особей, в качестве неизбежного следствия эволюции гиперциклов. Более детальный физико-химический анализ этого вопроса изложен в гл. 6. В своей книге он, как и я, рассматривает опыты С. Спигелмана в качестве иллюстрации верности теоретических критериев естественного отбора.
Эйген следующим образом подводит итоги всему сказанному: «Эволюция представляется неизбежным событием, если задано присутствие определенного вещества с определенными авто- каталитическими свойствами и если поддерживается такая величина потока (свободной) энергии, которая необходима для компенсирования стационарного производства энтропии» [350, с. 185]. «Тот факт, что «отбор» и «эволюция»—по аналогии с «равновесием» в термодинамике — можно охарактеризовать экстремальными принципами, позволяет физически обосновать и количественно сформулировать дарвиновскую теорию. В такой форме теория Дарвина уже не просто описывает некий исторический путь, но представляет собой выводимый из физики закон, управляющий общими процессами самоорганизации материи» [350, с. 197]. «Процессы автокаталитического отбора отфильтровывают и усиливают мутантов, обладающих высокой селективной ценностью, снижая тем самым недетерминированность, однако недетерминированность все-таки сохраняется в отношении выбора индивидуальных копий и затем отображается на макроскопическом уровне. Вследствие этого невозможно с достаточной точностью ни проследить прошлую, ни предсказать будущую эволюцию за пределами известных временных границ...» [350, с. 199].
При сопоставлении наших взглядов для дальнейшего изложения, вероятно, наиболее существенно сравнение понятий селективного преимущества и кинетического (биологического) совершенства. Смысл этих понятий очень близок. И и /; служат критериями в процессах естественного отбора. Величина Wm, введенная Эйгеном, аналогична понятию эволюционный потенциал данного направления эволюции, введенному выше.
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги