5.Кости, гены и фонемы
Ни Протш, ни Бомон не были палеоантропологами в собственном смысле слова: анализ ископаемых человеческих костей не входил в сферу их профессиональной компетенции. Основной упор в своих построениях они делали на имевшиеся в то время еще немногочисленные и не слишком надежные абсолютные датировки, на данные о геологических условиях залегания находок и на характер сопровождающих кости каменных орудий, но не на сопоставление скелетных остатков ранних сапиенсов между собой и с остатками других гоминид и поиск их вероятных предков. Первым, кто вплотную занялся разработкой теории африканского происхождения гомо сапиенс на основании именно палеоантропологических материалов стал немецкий исследователь Гюнтер Бройер. В серии статей, опубликованных в 1981-1984 гг., он проанализировал имевшиеся в то время костные материалы, определил их возможные эволюционные связи между собой и наметил основные стадии процесса формирования того круга признаков, который отличает анатомически современных людей. Это процесс, по мнению Бройера, совершался в Африке и не прослеживается на других континентах. В Европе параллельно с ним происходила неандертализация местного населения, а на большей части Азии сохранялись популяции потомков местных эректусов, которые еще не очень отличались от своих предшественников.
Вскоре с близкими моделями происхождения гомо сапиенс выступили и другие антропологи, такие как американец Филип Райтмайр и англичанин Кристофер Стрингер, что вызвало немедленную реакцию оппонентов, среди которых особенно активны были полицентристы, именовавшие теперь свои взгляды мультирегионализмом (Милфорд Уолпоф). С середины 80-х годов вопрос о времени и месте первоначального появления анатомически современных людей занял ведущее место на страницах многих научных изданий, стали регулярно проводиться международные конференции и издаваться сборники по этой теме, а дебаты между сторонниками и противниками африканской модели (то есть, фактически, между моно- и полицентристами) приобретали все больший накал. В 1990г., публикуя первый в отечественной литературе подробный обзор и разбор новых данных по происхождению гомо сапиенс, автор этих строк предположил, что, "возможно, впоследствии будет констатировано, что именно в конце 80-х годов дискуссия достигла апогея".12 К такому же выводу, но оценивая события уже в ретроспективе, пришли современные историографы, включая одного из самых активных участников тогдашних дебатов. 13
Баланс аргументов, которые могли привести в защиту своих взглядов сторонники противоборствующих точек зрения, менялся очень быстро. В середине 80-х годов все еще возможно было достаточно убедительно обосновать тезис, что анатомически современные люди появились в разных регионах Старого Света примерно в одно и то же время, где-то в интервале от 35 до 45 тысяч лет назад. Многим ученым по-прежнему казалось, что полицентристская модель лучше согласуется с фактами. Однако уже к концу десятилетия ситуация коренным образом изменилась. Появившиеся буквально в течение 3-4 лет новые абсолютные датировки ряда ключевых находок и генетические исследования решительно склонили чашу весов в пользу моноценристской модели. Важной вехой в развитии дискуссии стала опубликованная в 1988 г. в журнале "Science" статья Питера Эндрюса и Кристофера Стрингера "Генетические и ископаемые данные о происхождении современных людей". В ней были проанализированы и обобщены многочисленные факты, полученные генетиками, антропологами, археологами и специалистами по разным методам абсолютного датирования, и показано, что удовлетворительно объяснить их можно лишь с позиций теории африканского происхождения гомо сапиенс. За прошедшее с тех пор время - а миновало уже более четверти века - фактов, свидетельствующих в пользу этой теории, стало намного больше, а к числу дисциплин, выступающих в роли их поставщиков, добавилось несколько новых, прежде всего, лингвистика. Эти факты можно разделить на несколько основных групп.
1) Ископаемые палеоантропологические материалы однозначно свидетельствуют о том, что люди современного или очень близкого к таковому анатомического типа появились в Африке гораздо раньше, чем где бы то ни было еще. Если не считать юг Ближнего Востока, являющийся в географическом и тектоническом плане как бы продолжением Африки (Красное и Мертвое моря - это северная оконечность Великой рифтовой системы, простирающейся на юге до оз. Ньяса в Малави), то разница составляет не менее 100 тысяч лет.
Самой древней из известных сейчас антропологических находок, относимых к гомо сапиенс, является уже упоминавшийся череп Омо 1 с северного побережья оз. Туркана в Эфиопии. Его возраст, судя по имеющимся сейчас абсолютным датировкам и ряду других данных, составляет от 190 до 200 тыс. лет назад. Лобная и особенно затылочная кости этого черепа анатомически вполне современны, равно как и остатки костей лицевого скелета. На сохранившемся фрагменте нижней челюсти фиксируется достаточно развитый подбородочный выступ. По заключению многих антропологов, изучавших эту находку, череп Омо 1, а также известные части посткраниального скелета того же индивида не несут признаков, выходящих за рамки обычного для гомо сапиенс размаха изменчивости.
К находкам из Омо в целом очень близки по своему строению черепа, найденные в самом конце прошлого века на местонахождении Херто в Среднем Аваше, тоже в Эфиопии. Один из них, Херто 1 (рис. 5.1), дошёл до нас почти целиком (кроме нижней челюсти), другие представлены фрагментами. Возраст этих находок составляет от 154 до 160 тыс. лет. В целом, несмотря на наличие ряда примитивных признаков, морфология черепов из Херто позволяет рассматривать их обладателей как древних представителей современной формы человека. Объем мозговой полости черепа Херто 1 составляет 1450 см3, т.е. вполне типичен для нашего вида. При первоначальном описании в 2003 г. ископаемые из Херто были выделены в особый подвид гомо сапиенс - Homo sapiens idaltu, но насколько это оправдано пока неясно. Сопоставимые по возрасту черепа и другие кости людей современного анатомического типа были обнаружены и на ряде других восточноафриканских местонахождений, таких как Синга в Судане, грот Мумба в Танзании, пещера Дире-Дауа в Эфиопии. Таким образом, целый ряд хорошо изученных и довольно надёжно датированных антропологических находок из Восточной Африки свидетельствует о том, что люди, не отличавшиеся или мало чем отличавшиеся в анатомическом плане от нынешних обитателей Земли, жили в этом регионе уже 150-200 тыс. лет назад.
| Рис. 5.1. Череп раннего сапиенса с местонахождения Херто в Эфиопии (источник: Cartmill ans Smith 2009) |

В Южной Африке останки людей современного анатомического типа (зубы, нижние челюсти, фрагменты черепов и костей посткраниального скелета) возрастом от 70 до 100 тысяч лет были найдены при раскопках пещер в устье р. Класиес. Близкую древность могут иметь и относимые многими исследователями к гомо сапиенс антропологические находки из ряда других южноафриканских пещер (Бордер, ди Келдерс). В Северной Африке человеческие костные остатки, обнаруженные на таких археологических памятниках, как Джебел Ирхуд, Дар-эс-Солтан, Мугарет эль Алия (все Марокко) и Хауа Фтеа (Ливия), тоже принадлежат индивидам, приближающимся по многим важным признакам к людям современного анатомического типа (рис. 5.2). Судя по последним датировкам, возраст некоторых из них может достигать 160 тысяч лет. Анатомически полностью современный скелет ребенка, жившего примерно 50-60 тысяч лет назад, был найден на стоянке Тарамса в Египте.
| Рис. 5.2. Расположение упоминаемых в тексте африканских местонахождений скелетных остатков ранних сапиенсов и их непосредственных предшественников |

За пределами Африки пока не найдено останков гомо сапиенс, возраст которых превышал бы 45 тысяч лет. Единственным исключением является расположенный в непосредственном соседстве Ближний Восток, но об этом мы поговорим в одной из следующих глав, когда речь пойдет о путях и времени расселения нашего вида за пределы его прародины. Что касается остальной территории Евразии, то из всех известных здесь сейчас останков гомо сапиенс ближе всего к рубежу в 45 тысяч лет находится бедренная кость из Усть-Ишима в Омской области, случайно найденная в 2008 г. на берегу Иртыша. Возможна, она даже немного - на одну-две тысячи лет -древнее этой даты. Европейские материалы, по которым, главным образом, и судили раньше о древности гомо сапиенс, значительно уступают по возрасту перечисленным выше африканским ископаемым. То же самое можно сказать и о восточноазиатских материалах. Правда, в последнее время стали появляться сообщения, будто на юге Китая есть кости сапиенсов, почти не уступающие по возрасту африканским, но речь идет о старых находках, сделанных в ходе хозяйственных работ и лишенных точной привязки. Нет никакой уверенности ни в том, что морфология этих костей действительно является современной, ни в том, что опубликованные даты действительно отражают их возраст. Из достоверных находок сапиенсов в Китае самой древней является фрагментированный костяк из пещеры Тяньюань (недалеко от Чжоукоудяня). Его обладатель жил примерно 40 тысяч лет назад. Для восточной части Старого Света можно упомянуть еще почти целый череп гомо сапиенс из пещеры Ниа на о-ве Калимантан (Индонезия), возраст которого на основании серии радиоуглеродных дат оценивается в 40 тыс. лет, и скелет из погребения Манго 3 в Австралии, имеющий примерно такую же или даже чуть большую (до 43 тысяч лет назад) древность.
Таким образом, можно констатировать, что, если в Европе и Азии (за исключением Ближнего Востока) гомо сапиенс появляется, судя по имеющимся ископаемым материалам и абсолютным датировкам, не раньше или лишь немногим раньше 45 тысяч лет назад, то в Африке останки его древнейших представителей имеют возраст в четыре-пять раз больше.
2) Из всех континентов только в Африке известно достаточно большое количество останков гоминид переходного характера, позволяющих хотя бы в общих чертах проследить процесс трансформации местных гомо эректус в людей современного анатомического типа (рис. 5.2).
Особое место среди таких находок занимают два черепа, уже упоминавшиеся в предыдущих главах. Один из них был обнаружен в районе Флорисбада, а второй - в долине реки Омо (череп Омо 2). Оба они интересны и важны для нас, прежде всего, тем, что представляют собой останки существ, которым оставалось сделать буквально полшага, чтобы стать неотличимыми от современных людей в анатомическом отношении. Эти полшага были сделаны, вероятно, уже ближайшими их потомками, которые приобрели чуть более высокий лоб, в то время как их кости стали чуть менее массивными. Череп, очень похожий на флорисбадский и Омо 2 и притом прекрасно сохранившийся, был найден в 1980 г. в отложениях формации Нгалоба на местонахождении Лэтоли в Танзании. По большинству признаков он также выглядит почти современным, и лишь некоторая покатость лба и чрезмерно развитые надглазничные дуги заставляют отделять его обладателя от собственно гомо сапиенс. Эту же переходную стадию эволюции человека характеризует черепа из Илерета в Кении, Эяси в танзании и некоторые другие находки, датируемые временем примерно от 150 до 300 тысяч лет назад. Более архаичны останки человека с замбийского местонахождения Брокен-Хилл, часто также именуемого Кабве (рис. 5.3). Череп и другие фрагменты скелета, обнаруженные здесь еще в 1921 г., явно древнее костей из Флорисбада, Омо, Нгалобы и им подобных: их возраст, судя по геологическим данным, составляет не менее 300 тысяч лет и может достигать 600 тысяч лет. Примерно такую же древность и близкое строение имеют черепа из Ндуту в Танзании, Бодо в Эфиопии (рис. 5.4) и ряд других. По своему облику они больше еще напоминают эректусов, чем современных людей.
| Рис. 5.3. Череп из Кабве (Брокен-Хилл). Когда-то его называли африканским неандертальцем, а сейчас либо относят к виду человека гейдельбергского, либо зачисляют в особый вид человека родезийского (источник: Cartmill ans Smith 2009) |

| Рис. 5.5. Череп из Бодо - уже не эректус, но еще далеко не сапиенс (источник: Cartmill ans Smith 2009) |
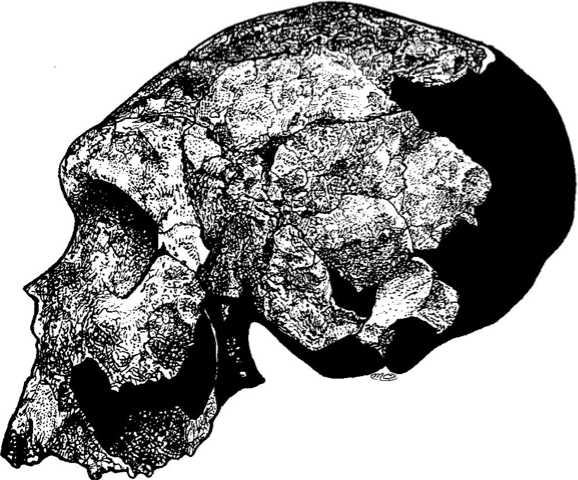
Всех африканских гоминид, анатомически и хронологически промежуточных между гомо эректус и гомо сапиенс, иногда относят вместе с их европейскими и азиатскими современниками к гомо гейдельбергенсис, а иногда зачисляют в особые виды, более ранний из которых именуется гомо родезиенсис (Homo rhodesiensis), а более поздний гомо гельмей (Homo helmei). Эти виды рассматриваются как последовательные звенья эволюционной цепочки, соединяющей последних эректусов с первыми сапиенсами (рис. 5.5). Ни в одном другом регионе Земного шара, кроме Африки, подобная цепочка пока не прослеживается.
| Рис. 5.5. Звенья эволюционной линии, идущей от эректусов к сапиенсам.1 - Бодо, 2 - Броккен-Хилл, 3 -Лэтоли, 4 - Омо 1, 5 - Бордер. |

3) Довольно убедительные аргументы в пользу африканского или. во всяком случае, южного, тропического происхождения первых европейских и ближневосточных гомо сапиенс были получены в результате сопоставления пропорций костей скелета у разных групп ископаемых и современных людей.
Известно, что человеческие популяции, издавна обитающие в районах с холодным климатом, — например, саамы Кольского полуострова, или эскимосы Аляски — отличаются укороченными по отношению к длине туловища конечностями, широким тазом, сравнительно большой для их роста массой тела и утолщенными трубчатыми костями. Как и следовало ожидать, все эти признаки наблюдаются и у неандертальцев, эволюционное становление которых совершалось в условиях приледниковой Европы. Напротив, первые люди современного физического типа и на Ближнем Востоке и в Европе по всем перечисленным параметрам демонстрируют сходство с коренными обитателями Африки и Австралии, т.е. регионов с жарким климатом. Как и последние, они характеризуются удлиненными конечностями, узким тазом, сравнительно небольшой по отношению к росту массой тела и более тонкими костями. Это явно свидетельствует против местного происхождения первых европейских гомо сапиенс. Интересно, что уже после исчезновения неандертальцев, в период интенсивного оледенения и похолодания, начавшийся примерно 25 тысяч лет назад и продолжавшийся не менее 10 тысяч лет, у обитателей Европы вновь начинают появляться многие особенности скелета, ранее свойственные неандертальцам. Предполагается, что это произошло вследствие приспособления к жизни в аналогичных природных условиях.
4) Данные генетики, по мнению подавляющего большинства специалистов в этой области, тоже указывают на Африку как наиболее вероятный первоначальный центр формирования вида гомо сапиенс. Наибольшее генетическое разнообразие среди современных человеческих популяций наблюдается именно там, а по мере удаления от Африки это разнообразие всё более убывает. Так и должно быть, если теория «африканского исхода» верна: ведь популяции гомо сапиенс, первыми оставившие свою прародину и обосновавшиеся где-то по соседству с ней, «захватили» в путь только часть видового генофонда, те группы, что затем отпочковались от них и продвинулись ещё дальше - только часть части и так далее.
С начала 80-х годов стало появляться все больше работ, в которых сравнивались последовательности нуклеотидов в митохондриальных ДНК разных групп приматов, в том числе и людей. Было показано, сначала на небольшом количестве выборок из разных расовых групп, что различия в этих последовательностях у людей очень малы - гораздо меньше, чем у других видов животных. Это было истолковано американским генетиком Уолтером Брауном как свидетельство того, что в эволюционной истории человечества сравнительно недавно имело место явление, называемое генетиками эффектом «бутылочного горлышка». Суть его состоит в том, что если в какой-то период существования данного вида численность его представителей по тем или иным причинам резко сокращается до размеров небольшой популяции, и если затем эта популяция все же не вымирает, а, наоборот, растет количественно, что приводит к восстановлению вида, то для последнего очень долго будет характерна очень низкая степень генетического полиморфизма, близкая к той, какая была у популяции-основателя. Браун не только обосновал вероятность прохождения человечества через подобное бутылочное горлышко, но и попытался определить древность этого события. По его расчетам оно случилось в промежутке от 360 до 180 тыс. лет назад.
Вскоре после этого по меньшей мере тремя группами ученых в разных лабораториях Японии и США было установлено, что в строении ДНК митохондрий современное африканское население обнаруживает гораздо большую степень вариабельности, чем население остальных континентов. Это логичнее и проще всего объяснить тем, что группа выходцев из Африки стала некогда предковой для всех нынешних жителей Земли, исключая лишь сам «черный континент», обитатели которого уже были к тому времени достаточно полиморфны в генетическом плане. Биохимики из Беркли (Р. Кэн, М. Стоункинг, А. Уилсон) рассчитали и вероятное время миграции из Африки группы, ставшей тем самым «горлышком», из которого «вылилось» все современное население Европы, Азии и Нового Света - 100 тыс. лет назад или несколько раньше. Как полагали тогда авторы этого исследования, мигранты на вновь осваиваемых ими территориях очень мало смешивались (если смешивались вообще) с предшествовавшим им автохтонным населением.
Работа Ребекки Кэн с соавторами появилась в печати в 1987 г., а годом раньше с обоснованием африканского происхождения современного человечества выступила другая группа исследователей, которые изучали полиморфизм не митохондриальной, а ядерной ДНК. «Анализ генетических дистанций, основанный на полиморфизме ядерных ДНК, - заключили Джон Уэйнкоут и его соавторы - показывает, что главная разграничительная линия проходит между африканским и евразийским населением», и имеющиеся данные соответствуют гипотезе о происхождении всех неафриканских популяций от популяции-основателя, мигрировавшей из Африки около 100 тыс. лет назад.
Впоследствии вывод о том, что основным (хотя, как мы еще увидим, вероятно, не единственным) источником генофонда всего современного населения Земли является именно Африка, был многократно подтвержден в работах других исследователей. Данные генетики человека, как и палеоантропологические данные, совершенно определенно указывают на Африку как исходный центр формирования гомо сапиенс.
5) Паразиты, досаждавшие человеку все время его существования, являются самыми "преданными" его спутниками, и не воспользоваться этим обстоятельством генетики, конечно, не могли. Была изучена изменчивость
ДНК ряда питающихся нами организмов, в частности, малярийного комара Plasmodium falciparum и бактерии Helicobacter pylori, паразитирующей на слизистой оболочке желудка и могущей вызвать язву желудка и двенадцатиперстной кишки. Географическая конфигурация генетической вариабельности в обоих случаях оказалась похожей на таковую у человека: по мере удаления от Африки эта вариабельность постепенно уменьшается, достигая своего минимума на окраинах ойкумены, заселявшихся позже всего (арктическая часть Европы, обе Америки, Полинезия).
5) Данные лингвистики, как стало выясняться в последние годы, дают ту же картину, что и генетические данные. Первым к этому выводу, насколько мне известно, пришел американец Пэтрик Мэннинг. Он проанализировал данные по классификации языков и количественному составу основных языковых семей и групп показал, что их современное географическое распространение наилучшим образом соответствует модели "африканского исхода".14 Через пять лет к точно такому же выводу, но совершенно другим путем, пришел новозеландский исследователь Квентин Эткинсон. В статье, опубликованной в журнале "Science", он убедительно продемонстрировал, что максимальное количество и разнообразие фонем существует именно в Африке, а по мере удаления от нее и первое, и второе уменьшается, достигая минимума в Южной Америке и Океании (рис. 5.6).
Рис. 5.6. Уменьшение фонетического разнообразия человеческих языков по мере удаления от Африки (источник: Atckinson 2011)
Канал с обзорами, анонсами новинок и книжными подборками
 Книжный Вестник
Книжный Вестник
Бот для удобного поиска книг (если не нашлось на сайте)
 Поиск книг
Поиск книг
Свежие любовные романы в удобных форматах
 Любовные романы
Любовные романы
О психологии, саморазвитии и личностном росте
 Саморазвитие
Саморазвитие
Детективы и триллеры, все новинки
 Детективы
Детективы
Фантастика и фэнтези, все новинки
 Фантастика
Фантастика
Отборные классические книги
 Классика
Классика
 ВКОНТАКТЕ
ВКОНТАКТЕБиблиотека с любовными романами, которая наверняка придётся по вкусу женской части аудитории
Любовные романы
Библиотека с фантастикой и фэнтези, а также смежных жанров
Фантастика
Самые популярные книги в формате фб2
Топ фб2
книги